

Introduction
Materials and Methods
Plant Materials and Growth Conditions
Shoot Fresh Weight
Total Phenolic Content and Antioxidant Capacity
Protective Effect of Cytotoxicity in Mugwort
Sugar Analysis in Kale
Statistical Analysis
Results
Plant Growth and Sugar Analysis
Total Phenolic Content and Antioxidant Capacity
Protective Effect of Cytotoxicity of Mugwort
Discussion
Plant Growth and Sugar Content
Total Phenolic Content and Antioxidant Capacity
Protective Effect of Cytotoxicity in Mugwort
Conclusion
Introduction
Plants require energy to complete their life cycle and produce sugars as an energy source during photosynthesis. Synthesized sugars are used as fundamental elements of plant growth and development via processes such as transport, utilization, and storage (Ruan, 2014). Representative sugars in plants
are glucose (glc) and fructose (fru), which are monosaccharides, and sucrose (suc), which is a disaccharide. Suc is a nonreducing sugar that consists of glc and fru. It has a limited chemical reactivity with other substances and can be easily metabolized to the sink in plants. Because of this advantage, suc is utilized as a major transport carbohydrate in plants (Mun and Yu, 2015).
Plants are exposed to continuously changing environmental conditions (i.e., response to stress). Using these sugars, plants stimulate the expression of genes involved in the adaptation and defense against stressful conditions as well as growth and development. This process is called “sugar signaling.” There is specific signaling of glc, fru, and suc in plants, which indicates that each type of signaling has different related enzymes, sugar sensors, and pathways. Therefore, plant responses and gene expression are different from each other (Ende and El-Esawe, 2014; Ruan, 2014). Suc-specific signaling is involved in the stimulation of fructan and proline synthesis, down-regulation of photosynthetic-related genes, and activation of anthocyanin biosynthesis genes in response to biotic and abiotic stresses (Julia et al., 2010). Since suc is a major sugar transporter in plants, it can be directly identified as a signal in plants. However, suc is easily decomposed to monosaccharides by hydrolysis, forming glc and fru. Thus, it is difficult to identify the pure effect of suc on the response of the plant (Tognetti et al., 2013). Several studies have utilized mutants and molecular approaches to identify the suc sensor, suc transporter, and pathway of suc signaling (Smeekens, 2000; Teng et al., 2005; Kühn and Grof, 2010; Shin et al., 2013). However, to date, the exact pathway or mechanism remains unknown (Shin et al., 2013).
Nevertheless, most previous studies have shown that suc treatment can express genes related to secondary metabolite biosynthesis (Hara et al., 2003; Hossain et al., 2009; Ferri et al., 2011; Guo et al., 2011; Zhang et al., 2013; Cao et al., 2014). When plants are treated with exogenous suc, it is absorbed by suc transporter 1, which is abundant in the roots of Arabidopsis and can produce signals to induce the expression of anthocyanin biosynthesis genes (Sivitz et al., 2008). In another study, explants obtained from lemon balm were cultured in Murashige and Skoog medium containing approximately 50-300 mM suc, which increased the contents of anthocyanin and flavonoid compounds along with a high level of suc (Hossain et al., 2009). The callus of grapevine also promotes phenylpropanoid synthesis in 40 g·L-1 suc treatment (Ferri et al., 2011). To date, the majority of studies related to exogenous suc were performed in tissue-cultured plants. Therefore, there is limited knowledge regarding suc signaling by suc absorbance via the roots in whole plants (Guo et al., 2011; Yang et al., 2012). Thus, the aim of this study was to validate the effect of exogenous suc on the enhancement of bioactive compounds using a hydroponic cultivation method to directly absorb suc with nutrients via roots in mugwort and kale plants.
Materials and Methods
Plant Materials and Growth Conditions
Mugwort (Artemisia princeps Pampanini cv. Sajabal) cuttings were prepared by cutting off stems, including the growing point, from mother plants. Rootone (ISK, Japan) was applied to the end of the cut stems. The cuttings were rooted in 105-cell seed trays containing growing medium (Myung-Moon; Dongbu Hannong, Seoul, Korea) in a growth chamber (DS-51GLP; Dasol Scientific, Hwaseong, Korea) under the following conditions: 22°C air temperature, 84% relative humidity, 150 µmol·m-2·s-1 photosynthetic photon flux density (PPFD), and 12-hour light period. After 17 days, growing medium from mugwort seedling roots was removed with flowing water. In total, 144 rooted mugwort plants were transplanted to deep-flow technique (DFT) systems (29.5 × 20.2 × 22.5 cm, L × W × H) with 6 holes. The plants were cultivated for 2 weeks under the following growth chamber conditions: 20°C air temperature, 73% relative humidity, 150 ± 5 µmol·m-2·s-1 PPFD, and 12-hour light period. A Hoagland nutrient solution (Hoagland and Arnon, 1950; pH 5.8 and electrical conductivity of 1.1 dS·m-1) was applied and replaced once before the suc treatments commenced.
Kale seeds (Brassica oleracea var. acephala ‘Manchoo Collard’) (Asia Seed, Seoul, Korea) were germinated in seed packs (Useem Instruments, Suwon, Korea), and distilled water was added once every 2 days. Kale seedlings were cultivated in a growth chamber under the same conditions as the mugwort seedlings for 2 weeks. After that, the same number of kale seedlings as for mugwort were transplanted into the DFT system and cultivated with Hoagland nutrient solution for 3 weeks before the suc treatments commenced. The nutrient solution was replaced every week.
After transplanting into each DFT system, 2-week-old mugwort and 3-week-old kale plants were subjected to Hoagland nutrient solution containing 10, 30, and 50 mM suc (D(+)-Sucrose; SAMCHUN, Pyeongtaek, Korea) for 4 and 3 days.
Shoot Fresh Weight
Shoot fresh weight of mugwort and kale plants was measured at 4 days and from 1 to 3 days of treatment, respectively. An electronic scale (Si-234; Denver Instrument, Bohemia, NY, USA) was used to measure the shoot fresh weights of both plants. Mugwort and kale samples were stored in a deep freezer (IU1786D; Thermo Fisher Scientific, Marietta, OH, USA) at -70°C until analysis of bioactive compounds.
Total Phenolic Content and Antioxidant Capacity
Fresh mugwort samples were extracted with 100 mL of 100% methanol in a shaker (Eyela Model MMS-300; Tokyo Rikakikai, Tokyo, Japan) at room temperature for 24 hours and the extract filtered through Toyo No. 2 filter paper (Toyo Ltd., Tokyo, Japan). To remove methanol, the filtrate was evaporated at 35°C. The dried extracts were dissolved in dimethyl sulfoxide (DMSO) at 50 mg·mL-1 and stored at -20°C until analysis. Total phenolic content was determined using the Folin-Ciocalteu method (Dewanto et al., 2002). Antioxidant capacity was estimated following the method described by Re et al. (1999) using 2,2'-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS; Sigma-Aldrich, St. Louis, MO, USA).
The analysis of total phenolic content and antioxidant capacity of kale leaves was performed by the Folin-Ciocalteu colorimetric method (Ainsworth and Gillespie, 2007) and the ABTS method (Miller and Rice-Evans, 1996) with slight modifications as follows. Frozen leaves (0.2 g) stored in the deep freezer were ground with liquid nitrogen using a mortar and pestle. The sample powder was mixed with 3 mL of 80% acetone and then transferred to a 2-mL microtube. For the analysis of total phenolic content and antioxidant capacity, the extract solution was incubated in a refrigerator at 4°C and a freezer at -20°C, respectively, in dark conditions for 12 hours. The analyses were performed based on the procedures described by Son and Oh (2013). Total phenolic content was expressed as milligrams of gallic acid equivalent (GAE) per gram of fresh weight, and antioxidant capacity was expressed as millimolar of trolox equivalent antioxidant capacity (TEAC) per gram of fresh weight.
Protective Effect of Cytotoxicity in Mugwort
Human hepatoma (HepG2) cells were provided by the Korean Collection for Type Cultures (Dejeon, Korea) and cultured in Dulbecco’s Modification of Eagle’s Medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco BRI, Gaithersburg, MD, USA), 100 U·mL-1 penicillin, 100 µg·mL-1 streptomycin, and 5.5 mM D-glucose (Sigma-Aldrich) in 5% humidity and carbon dioxide (CO2) at 37°C.
Cell viability was measured using the MTT assay (Mosmann, 1983). HepG2 cells were placed in 96-well plates at a density of 1 × 105 cells/well and incubated for 24 hours. The cells were then treated with 100 µg·mL-1 of the samples. After 24 hours, the cells were treated with ethanol (3%). After another 24 hours, 0.5 mg·mL-1 MTT reagent (Sigma-Aldrich), dissolved in FBS-free DMEM, was added to each well. After 2 hours of incubation, the culture medium was removed and the intracellular formazan product dissolved in DMSO (Sigma-Aldrich). The absorbance was measured at 550 nm using a spectrophotometer (Beckman Instruments Inc., Fullerton, CA, USA).
Sugar Analysis in Kale
Kale leaves were collected for sugar analysis at 1 and 3 days of treatment. The extraction method of Colaric et al. (2006) was applied with slight modifications as follows. A sample of 0.5 g of leaves was ground with 3 mL of 70°C deionized water using a homogenizer (PT 2500 E; Kinematica, Luzern, Switzerland) and incubated for 60 min at 70°C. The extracted samples were centrifuged at 15,000 × g for 10 min at 20°C. The supernatant was concentrated using a rotary evaporator (N-1000; EYELA, Tokyo, Japan) with a 60°C water bath, and then 1 mL of deionized water was added and sonicated using an ultrasonic sonicator (SK5210HP; Hangzhou Nade Scientific Instrument Co. Ltd., Zhejiang, China). The solution was filtered through a 0.45-µm UHP syringe filter (SPU4513-1; Woongki, Seoul, Korea). For the analysis of sugars such as fru, glc, and suc, a high-performance liquid chromatography system (YL9100, Young Lin Instrument Co., Ltd, Anyang, Korea) consisting of a YL9101 vacuum degasser, YL9101 quaternary pump, YL9131 column compartment, and autosampler was used. The separation of sugars was performed by a NH2P-50 4E column (4.6 × 250 mm; Shodex Asahipak, Showa Denko, Kanagawa, Japan). The column temperature was maintained at 35°C. Acetonitrile (75%) was used for the mobile phase with a flow rate of 0.5 mL·min-1. The total run time was 30 min, and a refractive index detector (Shodex Ri-101; Showa Denko, Kanagawa, Japan) was used to detect eluted sugars.
Statistical Analysis
We used a completely randomized block design. Five replications were performed for each mugwort treatment, except for the cell cytotoxicity test, which had three replications. For kale, six replications were performed for each treatment, except for sugar analysis, which had four replications. Analysis of variance was performed using a statistical analysis system (SAS 9.4; SAS Institute, Cary, NC, USA), and Duncan’s multiple range test was applied to compare means. Significant differences were considered at p < 0.05.
Results
Plant Growth and Sugar Analysis
To investigate the effect of nutrient solution containing different suc concentrations in a DFT system on the growth and suc content absorbed via roots, we measured the shoot growth in mugwort and kale plants and sugar content in kale leaves. Shoot fresh weights of suc treatments at 4 days in mugwort and 1 to 3 days in kale plants were not significantly different from the control (Table 1).
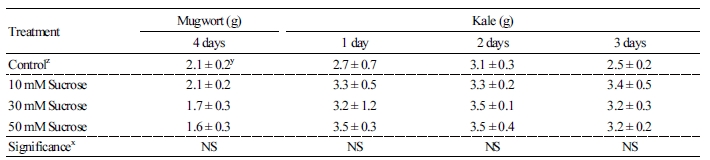
Sugar content (fru, glc, and suc) in kale leaves was higher than the control in all treatments (Fig. 1). At 1 day of treatment, the sugar content was higher in the order of 30, 50, and 10 mM suc treatments; however, there were no significant differences between treatments. The suc 50 mM treatment significantly induced 3.7-fold higher sugar content than the control at 3 days of treatment. In particular, suc content in kale leaves significantly increased 2.1-, 2.6-, and 2.2-fold in 10, 30, and 50 mM suc at 3 days of treatment, respectively, compared to the control.
Total Phenolic Content and Antioxidant Capacity
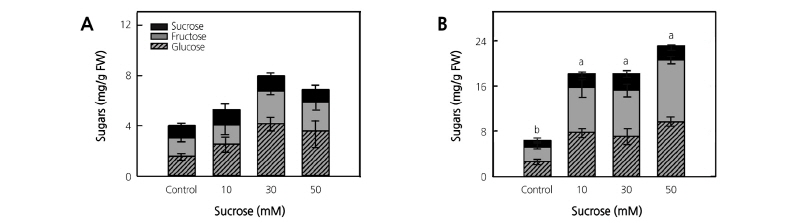
The suc-treated mugwort plants had increased total phenolic content and antioxidant capacity at 4 days of treatment (Fig. 2A and 2C). The total phenolic content of mugwort plants gradually increased 1.2-, 1.4-, and 2.3-fold in the 10, 30, and 50 mM suc treatments, respectively, compared to the control (Fig. 2A). In addition, the antioxidant capacity of mugwort plants gradually increased as suc concentration increased, with the 50 mM suc treatment having a 2.2-fold significantly higher antioxidant capacity than the control (Fig. 2C).
Fig. 2. Total phenolic content (A and B) and antioxidant capacity (C and D) of mugwort (A and C) and kale (B and D) plants treated with 10, 30, and 50 mM sucrose for 4 days at 2 weeks after transplanting (n = 5) and for 3 days at 3 weeks after transplanting (n = 6), respectively. The data indicate the means ± SE. Different letters above bars indicate significant difference at p < 0.05.
After 3 weeks of transplanting, the total phenolic content of suc-treated kale plants tended to increase with the treatment time and suc concentration for 3 days of treatment. At 3 days of treatment, there was no significant difference at the level of p < 0.05; however, total phenolic content was numerically 10~20% higher in the 10 and 30 mM suc treatments compared to the control (Fig. 2B). The suc 30 mM treatment at 1 day had 1.1-fold higher antioxidant capacity than the control, and the 10 and 30 mM suc treatments at 3 days had significantly higher (1.3- and 1.2-fold, respectively) antioxidant capacity than the control (Fig. 2D).
Protective Effect of Cytotoxicity of Mugwort
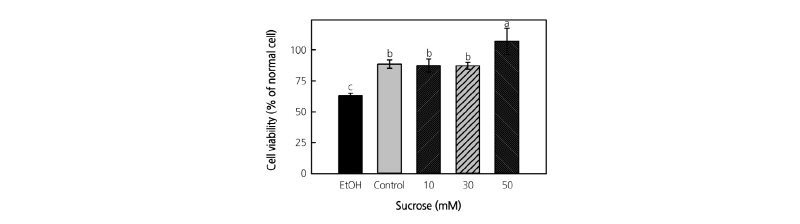
To investigate the protective effects of cytotoxicity of suc-treated mugwort plants, the extracts were pretreated with liver cells (HepG2 cells) damaged by EtOH. The suc-treated mugwort extract did not show any cytotoxicity (data not shown). The cell viability decreased to 63% after the EtOH treatment. However, the pretreatment of non suc-treated mugwort extract had significantly higher (1.4-fold) cell viability (88%) than EtOH-treated HepG2 cells. Mugwort extract treated with 10 and 30 mM suc did not have any effect on the cell viability. The only effect was observed in the 50 mM suc-treated mugwort extract with approximately 22% higher cell viability than the control and other treatments, which was close to the normal level of cell viability (Fig. 3).
Discussion
Plant Growth and Sugar Content
There are two ways for plant roots to uptake exogenous suc: (1) Suc can be hydrolyzed into monosaccharides (glc and fru) by cell wall invertase when placed in the environment around the root, and (2) suc can be directly taken up by the suc transporter that is located in the plasma membrane (Ruan, 2014; Veillet et al., 2016). Shin et al. (2013) reported that suc transporter 1 of Arabidopsis roots could be activated by increased transcript levels containing medium levels of suc (60 mM) and found that the absorption of suc was via the roots. Therefore, we expected that the exogenous suc treatments in nutrient solutions would also be absorbed by the suc transporter 1 of roots in the present study.
Callus from Vitis vinifera and explants from lemon balm plants grown in Murashige and Skoog media containing suc for 14 and 20 days, respectively, recorded significantly increased shoot fresh weights compared to the controls (Hossain et al., 2009; Ferri et al., 2011). However, in the present study, the exogenous suc supply via mugwort and kale plant roots did not influence the shoot fresh weights, even though suc-treated kale plants had significantly higher sugar content than the control. This result indicated that, unlike cell culture with photo-heterotrophs, 3 to 4 days of suc treatment is only a temporary treatment and does not affect the whole plant with photo-autotrophs (Yang et al., 2012).
Lobo et al. (2015) confirmed that exogenous 50 mM suc sprayed onto sugarcane leaves led to sugar accumulation in the attached leaves and triggered the down-regulation of Rubisco content and activity, resulting in severe reduction of net CO2 assimilation (PN) and instantaneous decrease of carboxylation efficiency (PN/Ci). Typically, suc concentrations of mesophyll cells are maintained below the threshold of suc concentration to prevent down-regulation of photosynthesis (Pollock et al., 2003). Although the threshold sugar content might be uncovered, a particular “beyond the threshold” content is required to generate appropriate signals in plants (Tognetti et al., 2013). In the present study, the suc content in kale leaves increased as suc treatment concentrations increased at 3 days of treatment. The 10, 30, and 50 mM suc treatments were 2.2-, 2.6-, and 2.2-fold higher, respectively, than the control. Since growth was not increased despite increased suc content, it can be expected that sugars accumulated above the threshold of down-regulation of photosynthesis in both mugwort and kale plants. Therefore, suc signaling induced these types of plant responses.
Total Phenolic Content and Antioxidant Capacity
In the present study, exogenous suc absorption via roots was effective to stimulate the biosynthesis of oxidative phenolic compounds. Ferri et al. (2011) confirmed that sugar content in grape callus tissue increased as medium suc concentrations increased. In addition, at 40 g·L-1 suc treatment, which was the highest level of sugar in the callus tissue, the highest levels of phenylalanine ammonia-lyase (PAL) and chalcone isomerase (CHI) enzyme activities were observed related to the phenylpropanoid biosynthetic process. Anthocyanin levels also increased. Zhang et al. (2013) reported that when hydroponically grown begonia seedlings were treated with 1.5 mM suc in Hoagland nutrient solution for 30 days, PAL, CHI, dihydroflavonol 4-reductase (DFR), and UDP-glucose anthocyanidin flavonoid glucosyltransferase (UFGT) enzyme activities significantly increased in the stem, and there was a 4.4-fold higher anthocyanin content than the control. Similar to our results, this result confirmed that the plant response was caused by suc signaling from the absorption of exogenous suc in nutrient solution via the roots.
Exogenous suc treatments can increase not only anthocyanin levels, but also flavonoid compounds of lemon balm, glucosinolates of broccoli sprouts, and phenylpropanoids of grape callus (Hossain et al., 2009; Ferri et al., 2011; Guo et al., 2011). Similarly, total phenolic content and antioxidant capacity of mugwort after 2 weeks of transplanting increased as suc concentration increased. In addition, after 3 weeks of transplanting, the antioxidant capacity of kale plants significantly increased in the 30 mM suc treatment at 1 and 3 days of treatment. These results imply that suc taken up by the suc transporter 1 of roots acts as a signaling molecule in the cytosol to induce the expression of genes related to the shikimate pathway to synthesize bioactive compounds, while simultaneously being used as a carbon source to manufacture compounds.
Protective Effect of Cytotoxicity in Mugwort
Mugwort contains catechin, which is a component of phenols and is used to cure hangovers and improve liver function by protecting liver cells from alcohol damage (Kim and Lee, 1998). In the present study, a 3% ethanol treatment was sufficient to decrease cell viability compared to normal cells (HepG2 cells); however, the non suc-treated mugwort extracts had 40.4% higher cell viability than the ethanol-treated cells. Moreover, increased bioactive compounds by suc-treated mugwort (50 mM suc treatment) showed the protective effects for ethanol-induced cytotoxicity. Similar results have been reported by Yang et al. (2012), who administered a treatment containing 6% suc in nutrient solution to perilla grown in a greenhouse. The perilla leaf extract (6% suc-treated perilla extract) had higher contents of phenolics, such as caffeic acid and rosmarinic acid, than the control and showed a significantly higher recovery of HepG2 cells than the cells that induced oxidative stress by tert-butyl hydroperoxide (t-BHP).
Conclusion
Since the majority of previous suc-related studies have been performed on tissue culture plants, the present study is valuable in that it demonstrated the improvement of bioactive compounds in whole plants that absorbed suc via their roots. In both mugwort and kale plants, the different suc treatments did not affect shoot growth; however, the sugar content in kale leaves increased by suc absorbed via roots. The increased patterns of antioxidants in mugwort and kale plants were different; however, our results identified that suc treatment stimulated the biosynthesis of antioxidants and thereby probably induced the protective effect of ethanol-induced cytotoxicity of suc-treated mugwort plants.
