서 언
재료 및 방법
실험 재료
생장조절제 처리
해부학적 관찰
과실의 수확시기 판정 및 품질 조사
통계분석
결과 및 고찰
생장조절제 처리에 따른 최종 수확량
생장조절제 처리에 따른 형태적 특성
생장조절제 처리에 따른 무핵률과 열과율
생장조절제에 따른 시기별 과실 품질
서 언
국립원예특작과학원에서 육종한 대립계 품종인 ‘Heukboseok’ 포도는 열과가 거의 없고, 착색이 우수하며 숙기가 빨라 ‘Kyoho’ 포도의 대체 품종으로 유망하다. 그러나 이러한 장점에도 불구하고 과육이 쉽게 물러지는 특성이 있어서 고품질 과실을 재배하기에 어려움이 따른다.
현재 농가에서 ‘Heukboseok’ 포도에 많이 이용하는 생장조절제 처리는 ‘Kyoho’ 포도에 준하여 처리되고 있거나 농가마다 처리시기와 농도를 무분별하게 살포하고 있을 뿐만 아니라 처리 목적 또한 불분명하여 생장조절제를 처리하였을 때 과실 품질과 특성에 대한 구명이 필요한 실정이다.
생장호르몬은 단백질의 수용체 및 신호전달, 화학적 합성, 효소의 활성 등 여러 가지 작용에 의해서 과실의 세포분열과 세포비대 등에 영향을 미친다(Taiz and Zeiger, 2013). 이와 같은 연구는 고품질 과실을 생산하여 농가의 소득을 높이기 위한 수단으로 농업에 많이 이용되어 왔다. Gibberellins은 여러 과종에서 착립률과 과실의 크기를 높여 생산량을 증대시킬 목적으로 사용되었고(Goldberg-Moeller et al., 2013; NeSmith 2002) 수분과 수정이 이루어지기 전에 gibberellins을 처리하여 단위결과 과실 생산에도 이용되었다(Atwell et al, 2003; Jeong et al., 1998; Lee et al., 2003; Stutte and Gage 1990). Phenylarea 물질을 함유하고 있는 thidiazuron (TDZ)은 전형적인 cytokinin 활성을 나타내며 N6-치환형 아데닌유도체에 의해서 세포증식을 유도하여 세포분열을 촉진하는 역할을 하는데 이는 내생 cytokinin의 활성보다 더 효과적인 세포분열을 일으키기 때문에 과실의 비대 및 발달에 이용되었다(Amarante et al., 2002; Greene 1995; Mok et al., 1987).
또한 생장호르몬의 효과를 극대화하기 위해서 두 가지 이상의 물질을 혼용하여 처리하는 기술도 많이 보고되었다. Bangerth and Schröder(1994)는 사과에서 GA에 cytokinin 활성물질인 CPPU를 혼용하여 결실률이 증가하였음을 보고하였고, Byun and Kim(1995)과 Lee et al.(2013)은 ‘Kyoho’ 포도에서 GA3와 TDZ 혼용처리 농도에 따른 열과발생 정도 및 과실의 품질에 대해 보고하여 ‘Kyoho’ 포도의 생장조절제 처리 기준을 확립하였다. 또한 Famiani et al.(2007)은 GA3와 TDZ 혼용처리는 숙기를 촉진시킨다고 보고하여 생장조절제에 의해 탄수화물의 활발한 이동을 예상할 수 있었다. Plessis(2008)는 포도에서 GA3와 CPPU처리는 cytokinin 활성물질에 의해서 과피의 두께가 두꺼워지고 이것은 곧 과육의 조직을 단단하게 한다고 보고였고, Zilkah et al.(1995)도 ‘Hass’ 아보카도에서 cytokinin의 활성으로 과육의 중심 부위가 강화된다고 보고하여 생장조절제 처리가 경도 향상에 영향을 준다는 것을 시사하였다.
따라서 본 연구에서는 신품종 ‘Heukboseok’ 포도에 생장조절제를 처리하여 숙성진행에 따른 과실특성을 관찰하고 과실의 형태적인 분석을 통해 연화 지연 효과를 구명하고자 한다.
재료 및 방법
실험 재료
경기도 천안시 입장면 소재의 포도원에서 재식 거리 3m × 5m로 재식된 4년생 ‘Heukboseok’ 포도를 대상으로 실험을 실시하였다. 1차 생장조절제 처리시기인 만개기에 목표 수량을 990m2 당 1,800kg으로 설정하기 위하여 꽃송이를 결과지당 1-2개를 남기고 나머지는 제거하였으며 만개 후 14일에 생장조절제 2차 처리 후 송이 다듬기를 통하여 한 과방당 과립수를 43알을 남겨두어 과방중을 500g으로 조절하였다.
생장조절제 처리
생장조절제는 GA3 3.1% 수용제(동부한농)와 thidiazuron 1% 수화제(N-phenyl-N’-1,2,3-hidiazol-5-yl urea, Bayer CropScience)를 사용하였다. 생장조절제 처리는 농가에서 처리하는 기준에 준하여 만개기(6월 8일)에 1차 처리하였고 만개 후 14일(6월 22일)에 2차 처리하였다. 생장조절제 처리 농도는 GA3 12.5mg・L-1,25mg・L-1, 50mg・L-1 단용처리 및 각각의 GA3 농도에 thidiazuron 2.5mg・L-1를혼용하는 처리로 나누어 실시하였으며 혼용처리는 1차 또는 2차에만 혼용처리하는 구와 1, 2차 모두 혼용하는 처리구로 나누어 실험하였다. 각 처리는 3반복으로 하였으며 반복당 1주로 하여 총 30주의 포도나무를 사용하였다.
해부학적 관찰
시료제작은 처리구당 무작위로 선발하여 과립의 적도부위를 채취하여 2.5% glutaraldehyde 용액에 1차 고정한 후 1% osmic acid 용액에 2차 고정하여 40-100% ethanol을 이용하여 순차적으로 탈수하였다. 탈수된 절편은 ethanol과 propylene oxide(1:1) 용액에 15분간 침지 후 100% propylene oxide 용액으로 유리화하였다. 유리화된 절편은 propylene oxide와 epon을 각각 2:1, 1:1로 섞은 용액에 1시간 이상 침지한 후 epon 용액에 하룻밤 동안 담가두었다. Epon 용액이 충분히 흡수된 절편을 다음날 DMP-30이 첨가된 epon을 이용하여 silicon mold에 넣고 40°C에서 4일간 경화시켜 block을 제작하였다. 완성된 block은 ultramicrotome (PT-X, RMC, USA)을 이용하여 1,000nm로 자른 후 0.5% periodic acid 용액, Schiff’s reagent, 1% sodium bisulfate에 각각 30분, 15분, 15분간 처리하여 염색하였다. 염색된 시료는 광학현미경(BX51, Olympus, Japan)을 이용하여 관찰한 후 세포사진을 촬영하여 과피두께, 과피층수 및 단위면적당 세포수를 조사하였다.
과실의 수확시기 판정 및 품질 조사
숙기 판정은 ‘Kyoho’ 포도 숙기 판정용 칼라차트(국립원예특작과학원)를 이용하여 숙기 판정표의 9 이상 되는 과실을 기준으로 관행 이전 수확인 만개 후 90일(9월 6일)에 1차 수확, 관행 수확기인 만개 후 100일(9월 16일)에 2차 수확하였으며 지연수확기인 만개 후 110일(9월 26일)에는 전량 수확하여 총 3회에 걸쳐 수확하였다.
수확한 과실의 과방중, 과립중 및 과립의 종경, 횡경을 측정하였으며, 경도는 과립의 적도부의 과피를 얇게 제거한 후 직경 2mm plunger가 장착된 물성 분석기를 이용하여 2mm・s-1 속도로 5mm 깊이까지 압축강도를 측정하였고 N으로 표기하였다.
당도는 디지털 굴절 당도계(Atago Corp., PR-32, Japan)를 이용하여 가용성고형물(SSC)을 측정하였으며, 산도는 과즙 10mL에 증류수 40mL를 혼합하여 희석한 후 0.1N NaOH를 가하여 pH가 8.1이 될 때까지 적정한 후 NaOH 사용량을 tartaric acid의 상당량으로 환산하였다. 안토시아닌 함량은 각 과방에서 10과립을 무작위로 선발하여 1.1cm의 cork borer를 이용하여 10개의 disk를 만든 후 0.1N HCl과 100% MeOH를 15:85(v:v)의 비율로 혼합한 침출액에 담근 뒤 12시간 암처리하여 추출하였다. 추출된 용액은 spectrophotometer (UV-1240, Shimadzu, Japan)를 이용하여 530nm파장에서 흡광도를 측정한 후 Siegelman and Hendricks(1958)의 방법으로 환산하였다.
열과는 전체 과방에 대하여 조사하였고 열과가 발생한 과방의 비를 계산하여 각 처리에 대한 열과 발생률을 산출하였다.
통계분석
통계분석은 수집된 데이터를 PASW statistics 18(SPSS Inc., USA) 프로그램을 이용하여 Duncan 다중검정(p = 0.05)으로 유의성을 분석하였다.
결과 및 고찰
Table 1. Cluster weight, berry weight and total yield of Heukboseok’ grapes after treatment with GA3 and TDZ. |
Treatment (mg・L-1) | Cluster weight (g) | Berry weight (g) | Total yield (kg per 990 m2) |
Full bloom | 14 days after full bloom |
Non-treated | 494.1 az | 10.7 ab | 1,647 (91.5%)y |
GA3 treatment | |
12.5 | 12.5 | 476.1 ab | 10.4 bc | 1,589 (88.3%) |
25 | 25 | 494.2 a | 10.5 abc | 1,807 (100.4%) |
50 | 50 | 446.8 c | 9.9 cd | 1,648 (91.6%) |
GA3 + TDZ combined treatment | |
12.5 + 2.5 | 12.5 | 446.0 c | 9.5 d | 1,431 (79.5%) |
12.5 | 12.5 + 2.5 | 494.7 a | 11.1 a | 1,904 (105.8%) |
12.5 + 2.5 | 12.5 + 2.5 | 476.8 ab | 10.5 abc | 1,547 (85.9%) |
25 + 2.5 | 25 | 449.3 bc | 9.3 cd | 1,518 (84.3%) |
25 | 25 + 2.5 | 467.3 abc | 10.4 bc | 1,720 (95.6%) |
25 + 2.5 | 25 + 2.5 | 476.9 ab | 10.7 ab | 1,688 (93.8%) |
zDifferent lowercase letters within columns denote significant differences by Duncan’s multiple range teat, 5% level. yTotal yield compared to target yield. |
생장조절제 처리에 따른 최종 수확량
과방중을 500g으로 하여 목표 생산량을 990m2당 1,800kg으로 하였을 때 최종 생산량은 무처리구 91.5%, GA3 단용처리구 88.3%-100.4%, GA3와 TDZ 혼용처리구는 79.5%-105.8%로 생산되었다. 특히 TDZ 혼용처리 시 TDZ를 1차에 혼용처리할 경우 작은 과립이 생산됨으로써 과방의 무게도 작아 최종 수확량이 적어지는 것으로 조사되었으며 GA3 단용처리 시 12.5mg・L-1 농도의 수확량 감소는 2차 처리에 TDZ를 혼용처리할 경우 과립이 11.1g로 커지고 최종수확량도 105.8%로 목표 수확량을 초과하여 수확량이 회복되는 것으로 조사되었다(Table 1). 이는 ‘Kyoho’ 포도에서 무처리구 또는 만개 후 14일의 2차 처리에만 GA3와 TDZ를 혼용한 처리구가 큰 과방중과 과립중을 생산하였다고 보고한 바와 일치하였다(Lee et al., 2003; Lee et al., 2013). 만개 후 14일인 2차에 GA3와 TDZ를 혼용한 처리구에서 과립중이 컸던 이유는 cytokinin이 auxin과 상호작용을 하며 과실의 세포분열과 세포발달에 영향을 미친다는 보고에서 알 수 있듯이(Taiz and Zeiger, 2013) 만개 후 14일의 TDZ 처리는 내생 auxin과 상호작용을 하여 세포분열을 더 연장시킨 것으로 생각되었다. 이는 토마토에서 만개기에 GA3를 처리할 경우 auxin의 발생이 억제되어 씨 없는 과실이 생산된 뒤 과육이 발달하는 시기에 다시 auxin이 검출된다는 보고(Atwell et al., 2003; Jong et al., 2009)에 의해 확인할 수 있었으며 ‘Heukboseok’ 포도의 경우에도 만개기에 GA3의 처리로 어느 정도 억제되었던 auxin이 이후 분비되어 TDZ와 상호작용 통해 세포분열을 연장했을 것으로 생각되었다. 특히 GA3 고농도 단용처리 및 1차에 TDZ를 혼용하여 1차 처리시 생장조절제의 농도가 높았던 경우에는 과립중이 매우 작았다. Coombe(1960)와 Jeong et al.(1998)은 생장조절제를 처리하여 종자를 제거시킨 과실에서 과립중이 작아지는 현상이 두드러지는데 이는 내생 gibberellins의 수준이 영향을 받기 때문이라고 하였다. 그러므로 본 연구에서 1차 처리시 GA3 고농도 처리나 TDZ 혼용처리는 내생 gibberellins 변화에 영향을 미쳐 과립 비대를 억제했을 것으로 생각되었다.
생장 조절제 처리에 따른 안토시아닌 함량 및 수확시기
Table 2. Anthocyanin concentration and comparison of yield after GA3 and TDZ treatment of ‘Heukboseok’ grapes. |
Treatment (mg・L-1) | | Harvest rate (%) | Anthocyanin (µg・cm-2) |
Full bloom | 14 days after full bloom | 1st 90 DAFBz | 2nd 100 DAFB | Final 110 DAFB |
Non-treated | 48.3 | 14.1 | 37.7 | 0.080 ay |
GA3 treatment | | | | |
12.5 | 12.5 | 12.4 | 13.0 | 74.6 | 0.068 ab |
25 | 25 | 5.9 | 9.1 | 85.0 | 0.078 a |
50 | 50 | 3.4 | 5.4 | 91.2 | 0.072 ab |
GA3 + TDZ combined treatment | | | | |
12.5 + 2.5 | 12.5 | 28.2 | 0 | 71.8 | 0.063 b |
12.5 | 12.5 + 2.5 | 21.2 | 25.8 | 53.0 | 0.075 ab |
12.5 + 2.5 | 12.5 + 2.5 | 31.0 | 4.9 | 64.1 | 0.071 ab |
25 + 2.5 | 25 | 13.6 | 7.6 | 78.8 | 0.068 ab |
25 | 25 + 2.5 | 16.4 | 19.9 | 63.7 | 0.082 a |
25 + 2.5 | 25 + 2.5 | 31.1 | 14.7 | 54.2 | 0.079 a |
zDAFB; days after full bloom. yDifferent lowercase letters within columns denote significant differences by Duncan’s multiple range test, 5% level. |
착색을 기준으로 수확기를 설정하였을 때 생장조절제 처리에 따른 만개 후 100일까지의 수확률은 무처리구가 62.4%로 가장 높았고 GA3 단용처리구는 모든 처리구에서 8.8%- 25.4%로 낮은 수확률을 보였으며 특히 GA3 50mg・L-1 처리구는 8.8%로 가장 낮은 수확률을 보여 GA3의 농도가 높아질수록 낮아지는 결과를 보였다(Table 2). 이는 Ben-Arie et al.(1996)이 감에서 GA3 처리가 숙기를 지연시켰다는 보고와 일치하였으며 Chun et al.(2012)이 ‘Campbell Early’ 포도에서 GA3가 저농도일 때 고농도에 비해 성숙이 촉진되었다는 보고와도 일치하였다. 그러나 생장조절제 처리구는 무처리구에 비해 숙기가 늦어졌지만 TDZ를 혼용할 경우 숙기 지연을 상쇄시켰으며 TDZ를 2차인 만개 후 14일에 처리한 처리구에서 더 효과적이었다. 이는 ‘Campbell Early’ 포도와 ‘Hayward’ 키위에서 TDZ를 GA3와 혼용처리 하였을 경우 안토시아닌 함량이 증가하며 숙기도 촉진되었다는 보고와 일치하였다(Famiani et al., 2007; Chun et al., 2012). 따라서 ‘Heukboseok’ 포도에 생장조절제를 처리할 경우 2차 또는 1, 2차 모두 TDZ를 혼용할 경우에 숙기 지연을 방지할 수 있을 것으로 생각되며 특히 1차에만 처리하는 것은 피해야 할 것으로 판단되었다.
안토시아닌 함량은 0.063-0.082µg・cm-2로 조사되었고 생장조절제 처리에 따른 안토시아닌 함량의 차이는 적었으나 TDZ를 1차에만 처리했을 때 안토시아닌 함량이 낮게 조사되어 TDZ를 만개기인 1차에만 처리하는 것은 착색을 불량하게 하는 것으로 나타났다(Table 2). 일반적으로 안토시아닌 함량은 변색기에 합성되어 숙성기간을 거치면서 당 함량 증가와 함께 축적되는데, TDZ 처리는 당도를 낮추고 산도를 높여 숙기를 지연시킴으로써 안토시아닌 함량도 낮아진다고 보고되었다(Coombe and McCarthy, 2000; Reynolds et al., 1992). 그러나 본 연구는 무처리와 함께 TDZ를 2차에 처리한 처리구에서 안토시아닌 함량과 수확률이 높게 조사되어 Famiani et al.(2007)와 Lee et al.(2013)의 보고와 일치하였다. 이는 cytokinin 활성 물질인 TDZ 처리에 의해 잎의 광합성 작용이 활발해져 과실로 영양분의 이동이 촉진되어(Taiz and Zeiger, 2013) 안토시아닌 함량 증진에 의한 착색촉진이 진행되었을 것으로 생각되었다.
Table 3. Characteristics of the skin and flesh at 90 days after full bloom of ‘Heukboseok’ grapes treated with GA3 and TDZ. |
Treatment (mg・L-1) | | Skin | Flesh cell number per area (5 cm2) |
Full bloom | 14 days after full bloom | Cell layer number | Thickness (µm) |
Non-treated | 9.4 az | 173.6 c | 14.9 b |
GA3 treatment |
12.5 | 12.5 | 9.3 a | 201.5 bc | 15.6 ab |
25 | 25 | 9.5 a | 259.7 a | 18.1 a |
50 | 50 | 9.1 a | 236.9 ab | 16.9 ab |
GA3 + TDZ combined treatment |
12.5 + 2.5 | 12.5 | - | - | - |
12.5 | 12.5 + 2.5 | 9.1 a | 176.3 c | 16.1 ab |
12.5 + 2.5 | 12.5 + 2.5 | 9.5 a | 246.9 a | 17.5 ab |
25 + 2.5 | 25 | 9.0 a | 235.1 ab | 18.6 a |
25 | 25 + 2.5 | 9.6 a | 262.3 a | 16.3 ab |
25 + 2.5 | 25 + 2.5 | 9.3 a | 261.3 a | 17.8 ab |
zDifferent lowercase letters within columns denote significant differences by Duncan’s multiple range test, 5% level. |
생장조절제 처리에 따른 형태적 특성
생장조절제 처리에 따른 과실 조직의 변화를 관찰하기 위해서 과피층수와 두께 및 과육의 밀도를 조사하였다. 과피는 맨 위 한층인 외표피층과 아표피층으로 구성되어있고 모든 처리구에서 9.0-9.6층으로 유의성이 인정되지 않았다. 하지만 무처리구와 생장조절제 처리구의 아표피층까지의 두께는 각각 173.6µm, 176.3-262.3µm로 조사되어 GA3와 TDZ에 의해 세포비대의 효과로 과피가 두꺼워진 것으로 생각되었다. 반면에 과육의 밀도를 알아보기 위해 과육세포의 수를 측정한 결과 무처리구는 14.9개, 생장조절제 처리구는 15.6-18.1개로 생장조절제 처리구의 세포수가 많고 세포간극도 무처리구에 비해 나타나지 않아 단위 면적당 세포의 밀도가 높은 것으로 조사되었다(Table 3). 이는 Table 5에서 생장조절제 처리구의 경도가 높고 무처리구에 비해 숙성이 진행되는 동안에도 계속해서 높은 경도를 유지하는 결과와 상응되었다. 또한 아표피층 수는 변화가 없는데 세포비대에 의해서 과피가 두꺼워지고 과육의 밀도는 세포분열에 의해서 증가하는 것으로 보아 세포분열과 세포비대가 독자적으로 조절되기 때문으로 생각되었다. Lee et al.(2013)과 Kwon (2013)은 ‘Kyoho’ 포도와 ‘Gaeryangmeoru’에 GA3 및 TDZ 처리시 세포비대에 의해 과피가 두꺼워진다고 보고하였으며 또한 Plessis(2008)와 Ng et al.(2013)은 포도와 사과에서 cytokinin 활성에 의해서 세포분열이 활발해져 밀도가 증가함에 따라 경도가 높아지고 과육의 연화도 저하된다고 보고하여 본 결과와 일치하였다.
생장조절제 처리에 따른 무핵률과 열과율
생장조절제 처리에 따른 평균 무핵률은 54.7%였으며 GA3 단용 처리시 47.8%, TDZ 혼용처리시 58.1%의 무핵률을 보여 TDZ를 혼용처리 하는 것이 GA3 단용처리보다 높은 무핵률을 보이는 것으로 조사되었다. 특히 GA3 12.5mg・L-1 처리구에서는 1, 2차 모두 TDZ를 혼용처리 할 경우 무핵률을 높일 수 있었으며 GA3 25mg・L-1 처리구는 TDZ 혼용 시기 및 회수에 관계없이 무핵률이 높아지는 것으로 조사되었다. 반면에 가장 높은 무핵률을 보인 1, 2차 GA3 25mg・L-1 + TDZ 2.5mg・L-1 처리구에서도 무핵률은 65.5%에 불과하였다(Table 4). 이는 ‘Kyoho’ 포도에서 만개기와 만개 후 10일에 GA3와 TDZ를 혼용처리한 것과 ‘Jarang’ 포도에서 만개기에 GA3를 처리한 처리구에서 90% 이상의 무핵과실을 생산했다는 보고와 일치하지 않았다(Jeong et al., 1998; Lee et al., 2012; Lee et al., 2013). 그러나 Jeong et al.(1998)은 ‘Campbell Early’ 포도에 만개기 GA3 처리가 7%의 낮은 무핵률을 나타내었다고 보고하였는데 이는 ‘Kyoho’ 포도에 비해 배주의 발달이 빨랐거나 GA3 처리에 의한 비정상적인 배낭 발달, 발아력 감소 또는 주심의 퇴화력에 의해서도 품종별로 차이가 나타날 수 있다고 보고하였다. 따라서 ‘Heukboseok’ 포도에서는 무핵과 생산을 위한 생장조절제 처리는 지양해야 할 것으로 판단되며 Jeong et al.(1998)과 Okamoto and Miura(2005)가 ‘Campbell Early’와 ‘Delaware’ 포도에서 보고한 바와 같이 ‘Heukboseok’ 포도에서도 80% 이상의 무핵과 생산이 가능한지에 대한 연구가 추후 더 필요한 것으로 생각되었다.
Table 4. Seedless rate and berry cracking rate for ‘Heukboseok’ grapes after treatment with GA3 and TDZ. |
Treatment (mg・L-1) | Seedless rate (%) | Fruit cracking rate (%) |
Full bloom | 14 days after full bloom |
Non-treated | - | 0.5 a |
GA3 treatment | | |
12.5 | 12.5 | 31.1 cz | 0.6 a |
25 | 25 | 55.7 ab | 0.5 a |
50 | 50 | 56.7 ab | 0.2 a |
GA3 + TDZ combined treatment | | |
12.5 + 2.5 | 12.5 | 54.4 ab | 0.4 a |
12.5 | 12.5 + 2.5 | 43.4 bc | 0.0 a |
12.5 + 2.5 | 12.5 + 2.5 | 60.2 a | 0.3 a |
25 + 2.5 | 25 | 63.0 a | 0.8 a |
25 | 25 +2.5 | 62.0 a | 0.9 a |
25 + 2.5 | 25 + 2.5 | 65.5 a | 0.2 a |
zDifferent lowercase letters within columns denote significant differences by Duncan’s multiple range test, 5% level. |
열과 발생 정도는 모든 처리구에서 0.0-0.9%로 발생이 미비하여 유의성이 인정되지 않아(Table 4), Lee et al.(2013)이 ‘Kyoho’ 포도에서 GA3의 농도가 높을수록 열과 발생률이 많아진다고 보고한 것과 Hur et al.(2012)이 ‘Heukgoosul’ 포도에서 50%-60% 만개했을 때 GA3 처리는 열과율을 높인다는 보고와는 일치하지 않았다. 열과의 원인으로는 비가 온 뒤 과도한 물의 흡수에 의해서 세포벽의 팽압 증가(Considine and Kriedemann, 1972; Yamamura and Naito, 1985)와 과실의 cuticle 또는 물관부의 특징과 기능에 의해서 발생한다고 보고되어왔다(Sekse, 1998). 또한 Demirsoy and Demirsoy(2004)는 sweet cherry에서 cuticle 층이 두꺼운 품종일수록 열과 발생이 미비하다고 보고하여 추후 ‘Heukboseok’ 포도의 생장조절제에 따른 형태학적 분석에 대한 연구가 더 필요할 것으로 생각되었다.
생장조절제에 따른 시기별 과실 품질
무처리구와 생장조절제 처리구의 가용성 고형물 함량은 큰 차이를 보이지 않았다. 반면에 만개 후 90일, 100일, 110일에 수확한 무처리 과실의 가용성 고형물 함량은 16.1, 16.5, 17.0°Brix로 시간이 갈수록 증가하는 경향이었으나 생장조절제 처리구의 평균 가용성 고형물 함량은 16.6, 16.7, 16.8°Brix로 수확시기에 따른 차이가 거의 없어 생장조절제 처리시 가용성 고형물 함량이 일찍 증가하는 것으로 조사되었다(Table 5). 이는 Ryou et al.(2009)이 블루베리에서 GA3 처리가 당도를 높였다는 보고와는 일치하지 않았지만 Yun et al. (2013)과 Lee et al.(2003)이 ‘Formosa’ 자두와 ‘Kyoho’ 포도에 GA3 단용처리와 GA3와 TDZ를 혼용처리 했을 경우 무처리구와 당도 차이가 나지 않았다고 보고한 것과 일치하였다. 특히, 생장조절제 처리 시 당이 일찍 축적됨에도 불구하고 초기 수확률이 낮아(Table 2) 당도가 안토시아닌 함량에 미치는 영향은 제한적인 것으로 생각되었다.
Table 5. Soluble solid content, acidity, SCC/acidity and firmness in ‘Heukboseok’ grape at 90, 100 and 110 days after full bloom after treatment with GA3 and TDZ. |
Treatment (mg・L-1) | Soluble solids content (°Brix) | Acidity (%) | SCCz/acidity | Firmness (N) |
Full bloom | 14 days after full bloom |
90 DAFBy |
Non-treated | 16.1 bcx | 0.61 b | 26.7 a | 24.1 bc |
GA3 treatment | | | | |
12.5 | 12.5 | 16.1 bc | 0.61 b | 26.7 a | 22.6 c |
25 | 25 | 16.0 c | 0.67 ab | 24.2 ab | 25.8 bc |
50 | 50 | 16.4 abc | 0.72 a | 23.2 b | 27.3 abc |
GA3 + TDZ combined treatment | | | | |
12.5 + 2.5 | 12.5 | 17.0 a | 0.63 b | 27.2 a | 25.7 bc |
12.5 | 12.5 + 2.5 | 16.8 abc | 0.64 b | 26.6 a | 24.8 bc |
12.5 + 2.5 | 12.5 + 2.5 | 16.3 abc | 0.65 b | 25.4 ab | 23.0 bc |
25 + 2.5 | 25 | 16.7 abc | 0.65 b | 26.1 ab | 28.8 ab |
25 | 25 + 2.5 | 16.9 ab | 0.62 b | 27.5 a | 24.2 bc |
25 + 2.5 | 25 + 2.5 | 17.0 a | 0.68 ab | 25.1 ab | 31.7 a |
100 DAFB |
Non-treated | 16.5 bc | 0.59 a | 28.6 b | 22.6 bc |
GA3 treatment | | | | |
12.5 | 12.5 | 16.8 abc | 0.58 a | 28.9 b | 19.8 c |
25 | 25 | 16.6 abc | 0.59 a | 28.2 b | 26.2 ab |
50 | 50 | 16.6 abc | 0.52 b | 32.6 a | 26.4 ab |
GA3 + TDZ combined treatment | | | | |
12.5 + 2.5 | 12.5 | - | - | - | - |
12.5 | 12.5 + 2.5 | 16.6 abc | 0.56 ab | 30.1 ab | 25.8 ab |
12.5 + 2.5 | 12.5 + 2.5 | 17.4 a | 0.60 a | 29.3 ab | 26.2 ab |
25 + 2.5 | 25 | 16.0 c | 0.52 b | 30.9 ab | 26.5 ab |
25 | 25 + 2.5 | 17.1 ab | 0.55 ab | 31.3 ab | 24.1 bc |
25 + 2.5 | 25 + 2.5 | 16.3 bc | 0.56 ab | 29.3 ab | 29.5 a |
110 DAFB |
Non-treated | 17.0 ab | 0.47 d | 37.0 a | 20.9 b |
GA3 treatment | | | | |
12.5 | 12.5 | 16.5 ab | 0.52 abc | 31.8 b | 26.6 a |
25 | 25 | 17.0 ab | 0.54 ab | 31.5 b | 25.3 ab |
50 | 50 | 16.2 b | 0.53 abc | 31.1 b | 24.8 ab |
GA3 + TDZ combined treatment | | | | |
12.5 + 2.5 | 12.5 | 16.9 ab | 0.53 abc | 32.1 b | 25.6 ab |
12.5 | 12.5 + 2.5 | 16.9 ab | 0.55 a | 30.7 b | 24.4 ab |
12.5 + 2.5 | 12.5 + 2.5 | 17.0 ab | 0.53 abc | 32.5 b | 23.5 ab |
25 + 2.5 | 25 | 16.5 ab | 0.50 cd | 33.2 b | 26.8 a |
25 | 25 + 2.5 | 17.3 a | 0.51 bc | 33.9 ab | 23.1 ab |
25 + 2.5 | 25 + 2.5 | 17.1 a | 0.53 abc | 32.6 b | 25.8 ab |
zSoluble solids content yDAFB; days after full bloom xDifferent lowercase letters within columns denote significant differences by Duncan’s multiple range test, 5% level. |
산 함량은 무처리구가 평균 0.56%, 생장조절제 처리구가 평균 0.57%로 큰 차이를 나타내지 않았으나 감소 시기는 무처리구에서는 만개 후 100일 후에 크게 낮아지는 것으로 조사되었으며 생장조절제 처리구에서는 만개 후 90일 이후에 현저히 낮아지는 것으로 조사되었다(Table 5). 이는 ‘Gaeryangmeoru’에서 GA3와 TDZ 혼용처리에서 TDZ 농도가 높을수록 산도가 높아진 결과와 일치하지 않았지만(Kwon et al., 2013), ‘Campbell Early’, ‘Kyoho’ 및 ‘Satsuma mandarin’에서 GA3와 TDZ 처리와 상관없이 산의 변화는 영향을 받지 않았다고 보고한 바와 일치하였다(Jeong et al., 1998; Kang et al., 2004). 따라서 ‘Heukboseok’ 포도에서의 생장조절제 처리는 일찍 당도를 높이고 산 함량을 감소시켜 당과 산을 기준으로 볼 때는 착색을 기준으로 수확하는 것과는 반대로 숙기를 촉진시키는 결과를 보였다.
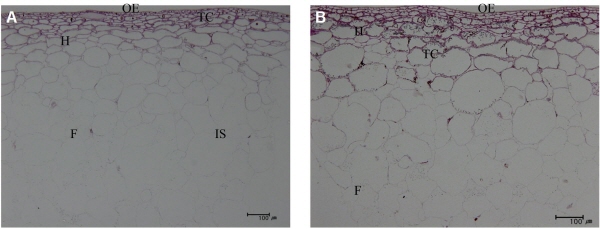
|
Fig. 1. Morphological structure at 90 days after full bloom of ‘Heukboseok’ grape treated with GA3 and TDZ. (A) non-treated; (B) primary and secondary 25 mg・L-1 GA3 + 2.5 mg・L-1 TDZ; F: flesh; H: Hypodermis; IS: Intercellular space; OE: Outer epidermis, TC: Tannin cell. |
|
 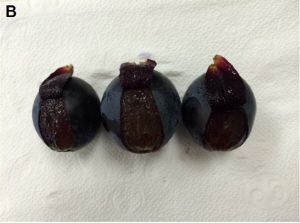
|
Fig. 2. Flesh appearance at the time of firmness measurement. (A) non-treated (B) Primary and secondary 25 mg・L-1 GA3 + 2.5 mg・L-1 TDZ. |
경도는 모든 수확시기에서 생장조절제 처리구가 무처리구보다 높게 유지되었으며, 만개 후 90일에서 110일 사이에 무처리구의 경도는 3.2N이 낮아졌으나 생장조절제 처리구는 0.9N이 낮아져 무처리시 과육의 연화가 촉진되었다(Table 5). 만개 후 90일과 100일에 GA3 25mg・L-1에 1, 2차 모두 TDZ를 혼용한 처리구에서 각각 31.7N, 29.5N으로 가장 높은 결과를 나타내었다. 동일한 시기의 무처리구의 경도는 각각 24.1N, 22.6N으로 낮게 조사되었으며 단용처리구와 혼용처리구는 처리간 뚜렷한 경향을 보이지 않았다. 그러나 과육의 밀도와 경도간에 수치적 차이가 약간 나는데 이는 1, 2차 GA3 25mg・L-1 + TDZ 2.5mg・L-1의 과피의 두께가 두꺼워 경도 측정 시 과피를 벗겨낼 때 과피의 가장 아랫부분이 과육에 남아있어 높은 경도가 측정되는 것으로 생각되었다(Fig. 2). 만개 후 110일에는 1, 2차 모두 GA3 25mg・L-1 + TDZ 2.5mg・L-1로 처리한 처리구도 만개 후 100일에 비하여 경도가 많이 낮아지는 결과를 보였다. 따라서 ‘Heukboseok’ 포도의 문제점인 쉽게 연화되는 현상은 Fig. 1과 Table 3에서 조사되었듯이 GA3와 TDZ 처리 시 과육의 밀도가 증가됨에 따라 개선됨을 확인하였고 특히 1, 2차에 모두 TDZ를 처리하였을 때 가장 효과적인 결과를 보여주었다. 하지만 ‘Hayward’ 키위와 ‘Formosa’ 자두에서는 GA3와 TDZ 처리시 경도가 낮아 과실의 연화가 빠르게 진행되어 숙기를 촉진하였다고 보고하여 본 연구와 상반되는 결과를 보고하였다(Famiani et al., 2007; Yun et al., 2013).
종합적으로 볼 때 ‘Heukboseok’ 포도는 생장조절제 처리에도 불구하고 무핵과실 생산이 어렵고 무처리구에 비해 착색이 빠르지 않아 생장조절제 처리시기에 따른 무핵과실 생산 및 생산량 조절을 통한 숙기 촉진은 어려울 것으로 판단되었다. 반면에 생장조절제 처리에 따른 가용성고형물 함량의 차이가 크지 않고 열과에 의한 2차 피해도 발생하지 않으며 과실연화를 지연시키는 효과가 있으므로 경도가 많이 낮아지기 전인 만개 후 100일 이전에 수확하기 위해서는 생장조절제 처리 농도를 1, 2차 모두 GA3 25mg・L-1 + TDZ 2.5mg・L-1로 처리하는 것이 과실 연화를 경감시키고 수확시기도 앞당길 수 있을 것으로 판단되었다.
Acknowledgements
본 연구는 농촌진흥청 공동연구사업(과제번호 : PJ008960122014)의 지원에 의해 수행되었음.
References
Amarante, C.V.T., P.R. Emani, L.E. Bassay, and C.A. Megguer. 2002. Thidiazuron effects on shoot growth, return bloom, fruit set and nutrition of apples. Pesq. Agropec. Bras. Brasilia. 37:1365-1371.
Atwell, B., P. Kriedenmann, and C. Turnbull. 2003. Plant in action. p. 361-363.
Bangerth, F. and M. Schröder. 1994. Strong synergistic effects of gibberellins with the synthetic cytokinin N-2-chloro-40pyridly)- N-phenylurea on parthenocarpic fruit set and some other fruit characteristics of apple. Plant Growth Regul. 15:293-302.
Ben-Arie, R., Y. Saks, L. Sonego, and A. Frank. 1996. Cell wall metabolism in gibberellins-treated persimmon fruits. Plant Growth Regul. 19:25-33.
Byun, J.K. and J.S. Kim. 1995. Effect of GA3 thidiazuron and ABA on fruit set quality of ‘Kyoho’ grapes. J. Kor. Soc. Hort. Sci. 36:231-239.
Chun, J.P., B.K. Kim, T.M. Bae, K.Y. Oh, and J.G. Kim. 2012. Effects of plant growth regulators sprayed at unfolded leaf stage on fruit quality in ‘Campbell Early’ grape. J. Agr. Life Sci. 46:9-15.
Considine, J.A. and P.E. Kriedemann. 1972. Fruit splitting in grapes: Determination of the critical turgor pressure. Aust. J. Agric. Res. 23:17-24.
Coombe, B.G. 1960. Relationship of growth and development to changes in sugars, auxins, and gibberellins in fruit of seeded and seedless varieties of vitis vinifera. Plant physiol. 35: 241-250.
Coomb, B.G. and M.G. McCarthy. 2000. Dynamics of grape berry growth and physiology of ripening. Aust. J. Grape Wine Res. 6:72-188.
Demirsoy, L. and H. Demirsoy. 2004. The epidermal characteristics of fruit skin of some sweet cherry cultivars in relation to fruit cracking. Pak. J. Bot. 36:725-731.
Famiani, F., P. Proietti, M. Pilli, A. Battistelli, and S. moscatello. 2007. Effcts of application of thidiazuron (TDZ), gibberellic acid (GA3), and 2,4-dichlorophenoxyacetic acid (2,4-D) on fruit size and quality of Actinidia deliciosa ‘Hayward’. N. Z. J. Crop Hortic. Sci. 35:341-347.
Goldberg-Moeller, R., L. Shalom, L. Shlizerman, S. Samuels, N. Zur, R. Ophir, E. Blumwald, and A. Sadka. 2013. Effects of gibberellins treatment during flowering induction period on global gene expression and the transcription of flowering control genes in Citrus buds. Plant Sci. 198:46-57.
Greene, D.W. 1995. Thidiazuron effects on fruit set, fruit quality, and return bloom of apples. HortScience. 30:1238-1240.
Hur, J.H., S.I. Oh, I.C. Son, Y.J. Oh, and D. Kim. 2012. Effect of GA3 treatment on the basis of period on berry cracking in ‘Heukgoosul’ grape. Kor. J. Hort. Sci. Technol. 30:133. (Abstr.)
Jeong, S.B., H.J. Lee, and S.J. Chung. 1998. Effect of acid on seedlessness induction and berry development in ‘Campbell Early’ and ‘Kyoho’ grapes by GA grown in Non-heated plastic house. J. Kor. Soc. Hor. Sci. 39:555-559.
Jong, M.D., C. Marlanl, and W.H. Vrlezen 2009. The role of auxin and gibberellins in tomato fruit set. J. Exp. Bot. 60:1523-1532.
Kang, S.B., C.W. Chae, S.H. Kim, J.W. Byun, K.J. Song, and D.K. Moon. 2004. Effect of foliar spray time of the mixture of GA3 and PDJ on the occurrence of peel puffiness and fruit quality in early-ripening Satsuma mandarin. Kor. J. Hort. Sci. Technol. 22:75. (Abstr.)
Kwon, Y.H., S.J. Kim, J.H. Choi, J.J. Choi, and H.S. Park. 2013. Gaeryangmeoru grapes response to GA3 and Thidiazuron treatment in the fruit anatomy and quality. Kor. J. Hort. Sci. technol. 31:121. (Abstr.)
Lee, B.H., Y.H. Kwon, Y.S. Park, and H.S. Park. 2013. Effect of GA3 an Thidiazuron on seedlessness and fruit quality of ‘Kyoho’ grapes. Kor. J. Hort. Sci. Technol. 31:135-140.
Lee, J.C., Y.L. Piao, J.K. Kim, K.S. Lee, and Y.S. Hwang. 2003. Induction of seedlessness in ‘Kyoho’ and Pione’ grapes (Vitis labruscana) with application of streptomycin, GA3, and Thidiazuron. J. Kor. Soc. Hort. Sci. 44:87-91.
Lee, J.W., J.H. Kim, E.Y. Hong, and Y.S. Lee. 2012. The effects of GA3 and Forchlorfenuron on the seedless berry ratio and fruit quality in ‘Jarang’ grape. Kor. J. Hort. Sci. Technol. 30:120-121. (Abstr.)
Mok, M.C., D.W.S. Mok, J.E. Turner, and C.V. Mujer. 1987. Biological and biochemical effects of cytokinin-activie phenylurea derivatives on tissue culture systems. HortScience 22:1194-1197.
NeSmith, D.S. 2002. Response of Rabbiteye blueberry (Vaccinium ashei Reade) to the growth regulators CPPU and gibberellic acid. HortScience 37:666-668.
Ng, J.K., R. Schröder, P.W. Sutherland, L.C. Hallett, M.I. Hall, and R. Prakash. 2013. Cell wall structures leading to cultivar differences in softening rate develop early during apple (Malus × domestica) fruit growth. BMC Plant Biol. 13:1471-2229.
Okamoto, G. and K. Miura. 2005. Effects of pre-bloom GA application on pollen tube growth in cv. Delaware grape pistils. Vitis. 44:157-159.
Plessis, B.W. 2008. Cellular factors that affect table grape berry firmness, p. 1-84. AgriSciences at Stellenbosch University.
Reynolds, A.G., D.A. Wardle, C. Zurowski, and N.E. Looney. 1992. Phenylureas CPPU and thidiazruon affect yield components, fruit composition, and storage potential of four seedless selections. J. Am. Soc. Hortic. Sci. 117:85-89.
Ryou, M.S., Y.Y. Hur, K.H. Kim, H.Y. Ju, and J.H. Roh. 2009. Influence of GA on the fruit set and Fruit quality of ‘Duke’ and ‘Darrow’ highbush blueberry cultivars grown in greenhouse. Kor. J. Hort. Sci. technol. 27:36. (Abstr.)
Sekse, L. 1998. Fruit cracking mechanisms in sweet cherries (Prunus Avium L.) - a review. Acta Hortic. (ISHS) 468:637-648.
Siegelman, H.W. and S.B. Hendricks. 1958. Photocontrol of Anthocyamn Synthesis in Apple Skin. Plant Physiol. 33:185-190.
Stutte, G.W. and J. Gage. 1990. Gibberellin inhibits fruit abscission following seed abortion in peach. J. Am. Soc. Hortic. Sci. 115:107-110.
Taiz, L. and E. Zeiger. 2013. Plant Physiol., p. 524-616. In: Pub. Sinauer (5th ed.).
Yamamura, H. and R. Naito. 1985. Susceptibility to berry splitting in several grape cultivars. J. Jpn. Soc. Hortic. Sci. 53:390-395.
Yun, S.K., H. Bae, I.K. Yoon, E.Y. Nam, J.H. Kwon, J.H. Jun, and K.H. Chung. 2013. Effect of GA paste on physiological fruit drop and fruit characteristics in ‘Formosa’ plums (Prunus salicina Lindl.) Protected Hort. Plant Fac. 22:309-315.
Zilkah, S., I. David, Y. Yeselson, M. Tamir, and L. Winer. 1995. Increasing ‘Hass’ avocado fruit size by CPPU and GA application, p. 11-18. In: Proceedings of The World Avocado Congress III.


