

서 언
재료 및 방법
실험설계 및 생육조사
토양 중 암모늄태 및 질산태 질소 분석
유리아미노산 분석
카테킨, 카페인 분석
통계처리
결과 및 고찰
질소함량 및 차광에 따른 광합성 능력 변화
질소함량 및 차광에 따른 엽색도 변화
질소함량 및 차광에 따른 토양 유기태 질소변화
질소함량 및 차광에 따른 찻잎 내 유리아미노산
질소함량 및 차광에 따른 찻잎 내 카테킨, 카페인 함량
서 언
유기농업에서는 화학비료를 사용하지 않고 가축분퇴비, 녹비, 작물잔사 등 여러 가지 유기물 자원을 이용하여 양분관리를 하는 것이 원칙이다(Cho et al., 2012). 작물재배 시 유기자재를 처리하게되면 유기자재내의 유기태질소가 토양중에 무기화 되는 정도는 탄질비에 기인하며, 탄질비가 낮을수록 질소의 무기화가 빠르게 진행되어 작물의 초기 생육을 촉진시킬 수 있다(Gale et al., 2006). 작물에 흡수된 질소는 황과 더불어 필수원소의 첫 번째 그룹으로 식물이 가장 많이 필요로 하는 무기원소로 아미노산과 핵산을 포함하는 많은 식물세포 성분을 구성하며 질소가 결핍되면 식물생장이 급속하게 저해된다(Gale et al., 2006). 이 때문에 대부분의 화학비료는 질소, 인, 칼륨과 같은 대량원소의 무기염을 포함함으로서 결핍시 발생하는 생장 저해를 억제하고 생육을 촉진시키는 작용을 한다(Gale et al., 2006). 반면 질소 과잉시 작물의 조직이 연화되어 도복과 병해충 피해 빈도가 증가하고, 토양부영양화, 용탈로 인한 수질오염 등의 문제를 야기함에 따라 작물별, 토양 유기물 함량 등을 고려한 적정 시비량 연구가 꾸준히 진행되었으며(Rabalais et al., 2002), 화학비료 이외의 질소공급원인 풋거름 작물, 가축분퇴비, 채종유박, 피마자박 등을 양분공급용 자원에 대한 연구가 진행되고 있다(Cho et al., 2012; Shim et al., 2020a). 2018년 말차 주생산지인 하동에서 차광 전 유박시비를 통하여 아미노산 함량을 높여 차의 감칠맛을 더하며 신초의 녹색도를 높여 고급 말차 생산이 가능하였으며, 2019년에는 기존 질소함량 높다 높은 유박사용에 따라 부분적으로 질소과잉에 따른 엽조직이 연약해지고 아미노산 함량 등이 증가함에 따라 차광이 시작하는 5월 무렵 차먼지응애 등의 응애류 발생 빈도가 높아지는 등의 문제 등이 관찰되었다(Shim et al., 2020b). 따라서 차광재배를 통한 말차의 품질 향상과 질소과잉에 따른 문제를 줄이기 위한 적정 질소 시비가 필요하며, 본 연구를 통해 고급말차 생산시 반드시 병행되어야하는 차광재배에 적합한 질소시비량을 알아보고자 한다.
재료 및 방법
실험설계 및 생육조사
직경 20cm, 높이 30cm 화분에 무비상토 약 1L를 채우고 건실한 차나무 재래종 종자 20립씩 2019년 5월 파종하여 온실에서 2020년 2월까지 생육시킨 유묘를 사용하였다. 격리된 환경을 조성하기 위해 569 × 363 × 255mm 플라스틱 상자에 원활한 뿌리 생육을 위해 식양토 약 2L를 넣고 그 위에 2년생 차나무가 식재된 3개의 화분을 올렸다. 유박의 질소함량에 따른 시비량은 0(N-0), 4.5(N-4.5), 9(N-9), 12(N-12) 및 18%(N-18)로 시비하였으며, 1주 단위로 생육조사를 실시하였다. 차광 재배시 질소함량이 말차 품질의 중요한 요소로 작용하기 때문에 비차광처리(non-shading, NS)와 95% 차광망을 덮은 차광처리(shading, S)으로 구분하였으며, 고급말차 생산시 20일 이상 차광을 진행하여 본 실험 역시 3주간 차광을 진행하였다. 비파괴적 분석법인 광합성 능력과 찻잎의 엽 색도를 측정하였다. 광합성 능력 측정은 휴대용엽록소형광측정기(FluorPen, FP110/D, PSI, Czech republic)를 이용하여 고정핀을 2–3번째 찻잎에 고정하여 30분간 부분 암처리 후, Fv(maximal variable fluorescence) / Fm(maximal fluorescence intensity), Pi_Abs, ABS·RC-1, DIo·RC-1 값을 통해 찻잎의 광합성 능력을 측정하였고, 동일 위치에 SPAD-502(Minolta, Japan)을 이용하여엽록소함량을 조사하였다. 찻잎의 엽 색도는 chroma meter (CR-400, Konica minolta Inc., Japan)를 이용하여 CIE(Commission internationale de L'eclairage proceedings) 체계인 L*, a*, b* 값을 획득하고 아래 공식을 이용하여 a*·b*-1 ratio를 조사하였다(Hwang at al., 2018).
토양 중 암모늄태 및 질산태 질소 분석
토양 건조시료 5g에 45mL의 2M KCl을 넣고 1시간 동안 진탕한 후 Whatman No. 2로 여과하여 침출액을 각각의 실험에 이용하였다. 암모늄태 질소(NH4-N)는 indophenol-blue 비색법(Searle, 1984)을 응용하여 측정하였다. 침출액 2mL에 완충용액 A(증류수 100mL에 6.7g trisodium citrate dihydrate, boric acid 3.4g, citric acid dihydrate 1.94g 녹임), 완충용액 B(증류수 100mL에 phenol 3.5g, sodium nitroprusside 0.04g 녹임), 완충용액 C(증류수 100mL에 NaOH 2g, dichloroisocyanuric acid 0.2g 녹임)를 각각 0.2mL 혼합한 후 37°C에 30분간 중탕시키고 UV/VIS Spectrophotometer(Optizen 3220UV, Mecasy Co., Ldt, Korea)를 이용하여 667nm의 파장에서 측정하였다(Searle, 1984). 질산태 질소(NO3-N)는 brucine법(Wolf, 1944)을 응용하여 측정하였다. 침출액 2mL에 5M NaCl 0.4mL, brucine-sulfanilic acid solution 0.5mL, 황산 10mL, 증류수 10mL를 넣고 혼합한 후 90°C에 20분간 중탕시킨 후 수냉으로 충분히 식힌 다음 UV/VIS spectrophotometer를 이용하여 410nm의 파장에서 측정하였다.
유리아미노산 분석
시료 0.1g에 증류수 10mL을 가한 후 3시간 동안 진탕시켜 10% 5-sulfosalicylic acid dihydrate 1mL을 첨가해 12시간 동안 5°C에 방치시켜 단백질을 침전·제거하고, 4,000rpm에서 15분간 원심분리 후 상등액을 취해 감압농축기로 농축하였다. 감압농축물에 0.2M, pH 2.2 lithium citrate loading buffer 5mL을 첨가하여 희석한 후 0.45µm membrane filter로 여과한 여액을 아미노산 분석기(Skyam S7130, Amino acid reagent organize, Germany)를 이용하여 분석하였다(Kim et al., 2018).
카테킨, 카페인 분석
시료 0.5g을 50% 에탄올 50mL을 첨가하여 1시간 30분 동안 초음파 처리하여 추출하였다. 추출물을 여과(Whatman No.4)하고 걸러진 여과액 40mL과 동량의 etyhl acetate를 가하여 분획 후 상층액을 취하여 감압농축하였다. 감압농축물은 메탄올 20mL 정용 후 PFFL 0.45µm 필터로 여과하고, 10µL를 취하여 TSK-ODS 80TM column(4.5 × 250mm, 5µm, phenomenex, California, USA)이 장착된 HPLC(Ultimate 3000, Dionex, USA)를 사용하였다. 이동상은 0.2% H3PO4(on water) 용액과 100% acetonitrile을 85:15(v/v)의 비율로 2분간 유지한 다음, 70:30(v/v)의 비율로 40분까지 흘려주었다. 42분까지는 0:100(v/v)으로 하여 흘려주고 85:15(v/v)의 비율로 47분까지 흘려주었다. 유속은 1.0mL/min, 검출기(PDA-3000, Dionex)의 파장은 240nm로 하였다. 분석물질별 retention time(RT)은 (–)-epigallocatechin(EGC) 5.2분, (–)-catechin(C) 6.8분, caffeine 7.9분, (–)-epicatechin(EC) 8.9분, (–)-epigallocatechin gallate(EGCG) 9.5분으로 설정하였으며(Bae et al., 2001; Kim et al., 2018), 각각의 표준품은 Sigma-Aldrich Chemical Co.(St. Louis, MO, USA)로부터 구입하였다.
통계처리
모든 분석결과는 SAS program(Ver. 9.3, Cary, NC. USA) 통해 3회 반복하여 측정한 평균값과 표준편차를 나타내었으며, PROC ANOVA 분산분석 및 Duncan's Multiple Range Test 사후검정을 실시하여 평균값 p < 0.05 유의수준에서 유의성을 비교하였다.
결과 및 고찰
질소함량 및 차광에 따른 광합성 능력 변화
질소시비 및 차광 1주차의 최대양자수득율[변동형광값(Fv) · 최대형광값(Fm)-1]은 비차광, 차광처리 모두 질소함량 9%(이하 N-9)에서 각각 0.79, 0.83으로 가장 높았으며, 질소함량 12%(이하 N-12)를 제외한 모든 차광처리가 비차광처리 보다 높은 최대양자수득율을 보였다. 처리 2주차 역시 비차광, 차광처리 모두 N-9에서 각각 0.75, 0.83로 가장 높았으며, 차광의 경우 N-4.5, N-9가 근사치를 보였다. 처리 3주차의 결과를 보면, 비차광은 N-9에서 0.78로 가장 높았으며, 차광은 N-4.5에서 0.82로 가장 높았으며 이후 감소하는 경향을 보였다. Pi_Abs는 photosystem II(광계 II)에 의해 흡수된 광자와 광합성계 내부의 전자수용체 환원정도를 나타내고, 일반적으로 최대양자수득율과 정의상관을 보인다(Stribet and Govindjee, 2011). 처리 1주차의 Pi_Abs는 비차광은 N-9에서 3.68로 가장 높았으며, 차광은 N-4.5에서 2.56로 가장 높았다. 처리 2주차는 비차광은 N-9에서 1.82, 차광은 N-4.5에서 5.22으로 가장 높았으며, 처리 3주차에는 비차광은 N-9에서 2.76, 차광은 N-4.5에서 3.85로 가장 높았다. Yoo et al.(2012)에 의하면 고추잎에서의 Pi_Abs는 한발스트레스 처리 4일 후 약 30% 이상 감소하였다. 본 연구에서 질소과잉시 토양 내 전기전도도(EC, electrical conductivity) 등이 증가하여 뿌리로부터 수분 흡수가 저해됨에 따라 처리 시간이 길어질수록 고농도의 질소시비에서 생육이 저조한 것으로 사려된다. 반응 중심비(ABS·RC-1)와 열로 인한 에너지 소실(DIo·RC-1)는 작물체가 스트레스 정도와 정의상관을 보이는데, 처리 1주차부터 3주차까지 차광의 경우 N-12에서 가장 높았으며, 비차광은 1–2주차는 N-12에서 가장 높았으며, 3주차에서는 N-0에서 가장 높았다. 엽록소함량을 나타내는 SPAD는 더 건강하고 녹색의 잎일수록 수치가 높으며, 붓꽃(Iris germanica)의 경우 질소 비율이 높을수록 영양 성장이 촉진되어 SPAD 수치가 높게나타났다(Zhao et al., 2021). 엽록소함량은 처리2주차에서 비차광 처리구 N-12에서 45.60, 차광 처리구 N-9에서 48.87로 가장 높았으며, 처리3주차는 비차광 처리구는 동일한 N-12에서 43.03, 차광 처리구는 N-4.5에서 46.40으로 가장 높았다(Table 1). 비차광 처리구의 경우 질소 함량이 9–12%까지 큰 영향을 받지 않고 광합성능력을 유지하는 반면, 차광 처리구의 경우 처리기간이 길어질수록 질소과잉 장애가 관찰되었다. 질소는 식물을 구성하는 무기영양원소 중 가장 많은 비율을 차지하여 수체 생장에 미치는 영향은 매우 크다(Ha et al., 2019). 따라서 질소시비량이 높아지면 작물의 잎과 줄기 등에 질산환원효소활성이 증가하여 생육이 촉진되나 질소 과잉시비시에는 오히려 생장이 감소되기도 한다는 결과와 일치하였다(Gojon ea al., 1991; Sonneveld and Wood, 2009).
Table 1.
Comparison of photosynthetic capacity of tea leaves after 2 and 3 weeks according to nitrogen fertilization and shading treatment
| Tretment | Fv·Fm-1 | Pi_Abs | ABS·RC-1 | DIo·RC-1 | SPAD | ||
|
Week (A) |
Shading (B) |
Nz content (C) | |||||
| 2 week | NS | 0.0% | 0.664 cy | 1.287 b | 1.802 c | 0.636 b | 24.00 e |
| 4.5% | 0.717 ab | 1.490 b | 1.979 bc | 0.561 bc | 29.67 d | ||
| 9.0% | 0.754 a | 1.818 a | 2.022 b | 0.495 c | 34.97 c | ||
| 12.0% | 0.680 bc | 1.022 c | 2.428 a | 0.810 a | 45.60 a | ||
| 18.0% | 0.681 bc | 1.337 b | 2.147 b | 0.613 b | 39.73 b | ||
| S | 0.0% | 0.792 b | 2.683 b | 1.840 b | 0.378 b | 33.00 d | |
| 4.5% | 0.826 a | 5.220 a | 1.511 c | 0.254 c | 36.90 cd | ||
| 9.0% | 0.833 a | 5.066 a | 1.477 c | 0.278 c | 48.87 a | ||
| 12.0% | 0.772 c | 1.775 c | 2.031 a | 0.462 a | 37.93 c | ||
| 18.0% | 0.789 b | 3.155 b | 1.861 b | 0.399 b | 42.03 b | ||
| 3 week | NS | 0.0% | 0.632 d | 0.561 d | 2.259 a | 0.833 a | 29.67 c |
| 4.5% | 0.733 b | 1.114 b | 1.888 bc | 0.591 bc | 30.43 c | ||
| 9.0% | 0.784 a | 2.761 a | 1.792 c | 0.418 d | 34.07 b | ||
| 12.0% | 0.691 c | 0.932 c | 1.975 b | 0.617 b | 43.03 a | ||
| 18.0% | 0.713 bc | 1.205 b | 1.876 bc | 0.539 c | 28.97 cd | ||
| S | 0.0% | 0.806 b | 3.744 a | 1.541 c | 0.270 c | 44.40 b | |
| 4.5% | 0.822 a | 3.851 a | 1.405 d | 0.246 cd | 46.40 a | ||
| 9.0% | 0.808 b | 3.129 b | 1.712 b | 0.330 b | 31.80 c | ||
| 12.0% | 0.733 c | 1.152 d | 2.268 a | 0.611 a | 24.43 d | ||
| 18.0% | 0.793 bc | 2.534 c | 1.516 c | 0.339 b | 31.70 c | ||
| A × B × C | Sampling time | –0.15NS | 0.03NS | 0.37** | 0.36** | –0.19NS | |
| Shade type | 0.72** | 0.57** | –0.34** | –0.55** | 0.26* | ||
| N fertilization | –0.02NS | –0.12NS | 0.09NS | 0.08NS | 0.15NS | ||
zN; nitrogen, NS; non-shading, S; shading, Fv (maximal variable fluorescence) / Fm (maximal fluorescence intensity), Pi_Abs; performance index for energy conservation from photons absorbed by PSII antenna, until the reduction of PSI acceptors, ABS·RC-1; average absorbed photon flux per PSII reaction center, DIo·RC-1; heat dissipation at time zero, per RC.
차나무의 질소시비와 차광에 따른 광합성능력간의 상관관계를 통계프로그램(SAS program)으로 분석한 결과, 먼저 질소시비에 따른 광합성능력간의 상관관계가 없는 것으로 나타났다. 조사시기와의 상관관계에서는 활성화된 반응 중심비(ABS·RC-1)와 열로 인한 에너지 소실(DIo·RC-1)에서 고도로 유의한 상관관계를 보였다. 그러나 차광처리에 상관관계에서는 SPAD를 제외한 모든 특성에서 고도로 유의한 상관관계를 보이며, 특히 Fv·Fm-1에서 0.72**으로 가장 높은 정의 상관 관계를 보였다. 이처럼 차나무의 질소시비와 차광에 따른 광합성능력간의 상관관계에서 차광처리에 따른 뚜렷한 차이가 나타났다.
질소함량 및 차광에 따른 엽색도 변화
가루녹차는 찻잎의 녹색도에 따라 등급이 결정되기 때문에 차광 재배시 무엇보다 엽색을 최우선으로 한다(Hwang et al., 2018). 처리 1주차의 a*·b*-1 ratio는 음(-)의 값이 클수록 녹색정도가 높고, 양(+)의 값이 클수록 적색정도가 높은데, 차광 처리구는 N-9에서 –65.37로 가장 녹색정도가 높았으며, 비차광 처리구는 N-18에서 –51.71로 가장 녹색정도가 높았다. 처리 2주차도 역시 차광 처리구 N-9에서 –70.61로 가장 녹색정도가 높았으며, 비차광 처리구는 N-12에서 –65.76으로 가장 녹색정도가 높았다. 처리 3주차는 차광 처리구 N-4.5에서 –67.67로 가장 높았으며, 비차광 처리구는 N-12에서 64.19로 가장 높았다(Table 2). 수종이 다른 진달래의 차광 처리별 잎색의 변화를 Munsell color chart를 통해 관능검사결과 역시 전광 조건에서 R과 Y chart의 색이 많은 반면 차광 수준이 높을수록 G, Y의 chart에 속하는 비율이 높아 녹색을 많이 띄고 있다고 하였으며(Lee et al., 2010), 일반적으로 차광정도가 클수록 a*·b*-1 ratio의 음의 값이 증가한다는 연구 결과 및 질소함량 증가에 따른 녹색도 증가 결과와 일치하였다(Shim et al., 2019, 2020b). 질소시비에 따른 엽색도변화에서 a*·b*-1 ratio 을 제외한 모든 특성에서 상관관계가 없는 것으로 나타났다. 조사시기와의 상관관계 역시 a*·b*-1 ratio에서 고도로 유의한 상관관계를 보였다. 차광처리에 따라 광합성능력과 엽색도에 대해 연차 간의 추가 연구가 이루어진다면, 고품질 말차생산을 위한 기초자료로 활용도 높을 것으로 사료된다.
Table 2.
Color change of tea leaves after 2 and 3 weeks according to nitrogen fertilization amount and shading treatment
| Tretment | L | a* | b* | a*/b* ratioy | Chroma | ||
|
Week (A) |
Shading (B) |
Nz content (C) | |||||
| 2 week | NS | 0.0% | 50.6 bx | –10.9 a | 38.5 a | 32.3 c | 37.2 ab |
| 4.5% | 52.6 a | –13.5 b | 36.7 a | 37.2 c | 39.2 a | ||
| 9.0% | 45.8 c | –15.7 d | 31.9 b | 48.3 b | 35.5 bc | ||
| 12.0% | 41.5 d | –14.8 c | 21.5 d | 65.8 a | 27.7 d | ||
| 18.0% | 45.6 c | –15.1 cd | 29.0 c | 52.1 b | 32.7 c | ||
| S | 0.0% | 40.8 bc | –16.7 c | 25.0 c | 62.4 b | 29.7 c | |
| 4.5% | 42.8 ab | –15.5 b | 24.9 c | 65.2 b | 29.4 c | ||
| 9.0% | 38.6 c | –13.7 a | 19.7 d | 70.6 a | 24.0 d | ||
| 12.0% | 44.9 a | –19.1 d | 38.4 a | 49.7 c | 42.9 a | ||
| 18.0% | 45.0 a | –17.4 c | 26.8 b | 65.0 b | 32.0 b | ||
| 3 week | NS | 0.0% | 47.9 b | –15.2 b | 30.5 c | 41.8 d | 33.6 bc |
| 4.5% | 54.5 a | –15.1 b | 39.0 a | 43.4 c | 42.2 a | ||
| 9.0% | 52.8 a | –15.4 b | 38.4 ab | 52.9 b | 40.2 a | ||
| 12.0% | 45.1 c | –14.5 a | 24.9 d | 64.2 a | 28.7 c | ||
| 18.0% | 46.3 c | –14.4 a | 33.2 b | 50.7 b | 36.2 b | ||
| S | 0.0% | 42.7 c | –16.6 c | 21.8 c | 62.2 b | 32.4 ab | |
| 4.5% | 41.2 c | –14.7 b | 24.0 bc | 67.7 a | 30.3 b | ||
| 9.0% | 46.2 a | –17.2 c | 28.8 a | 62.6 b | 33.5 a | ||
| 12.0% | 44.1 b | –15.5 bc | 22.1 c | 58.1 c | 26.5 c | ||
| 18.0% | 44.0 b | –12.1 a | 25.7 b | 59.0 c | 28.4 c | ||
| A × B × C | Sampling time | 0.09NS | –0.10NS | –0.03NS | 0.31** | –0.14NS | |
| Shade type | –0.47** | –0.39** | –0.42** | 0.58** | –0.29** | ||
| N fertilization | –0.17NS | –0.14NS | –0.16NS | 0.23* | –0.14NS | ||
질소함량 및 차광에 따른 토양 유기태 질소변화
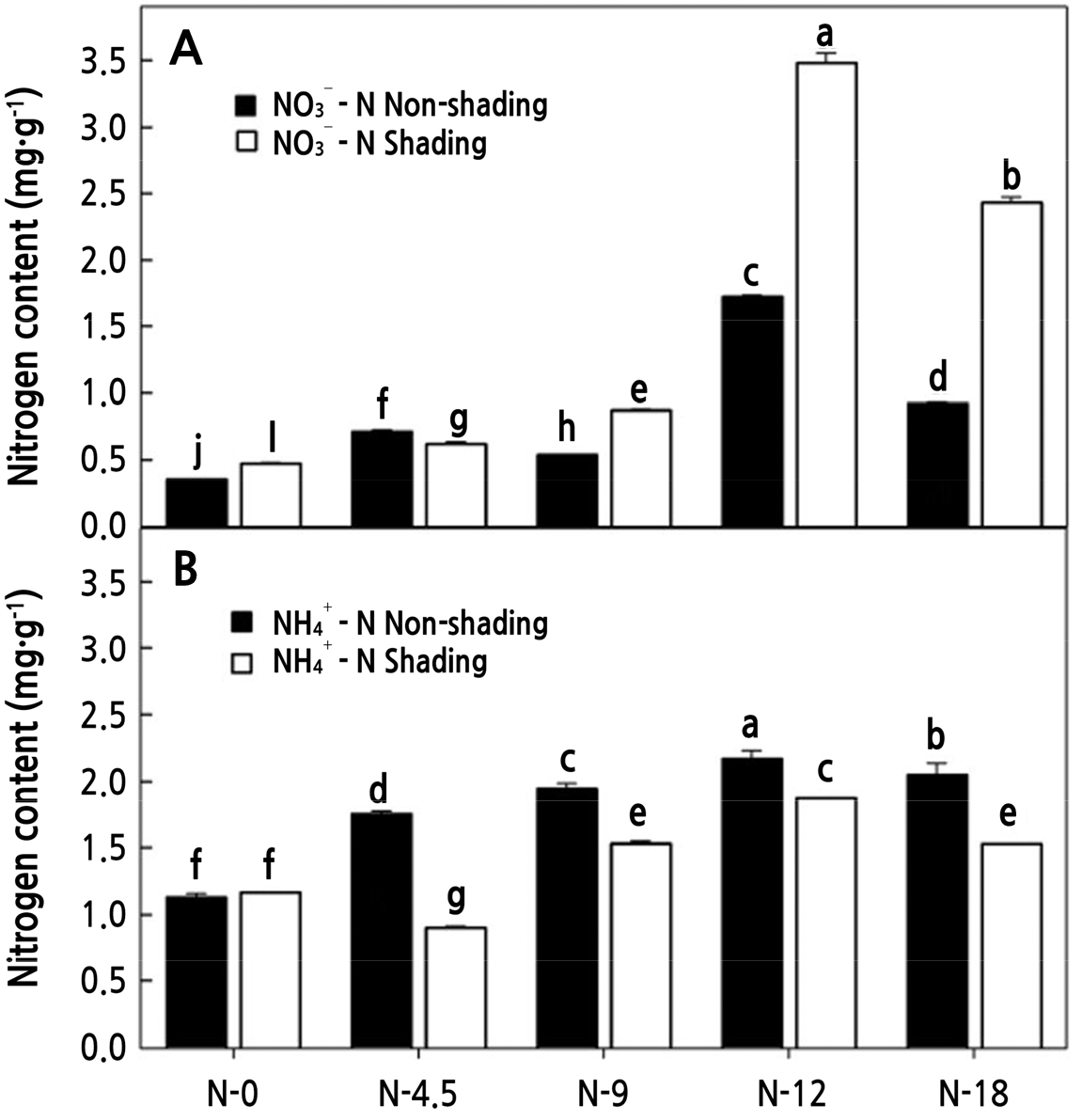
질소는 작물의 생육에 가장 큰 영향을 주는 원소로, 유박은 차나무의 대표적인 질소공급원으로 토양 중에서 암모늄태 질소로 분해된 다음 질산태 질소로 산화되면서 적합한 질소를 공급하게 된다(Vines and Wedding, 1960; Chapman, 1966; Kim et al., 2011). 질소 시비량에 따른 토양 내 유기태 질소 함량을 보면(Fig. 1), 암모늄태 질소는 차광조건하에서 N-12까지 점차적으로 증가하다가 N-18에서 감소하며, 비차광조건에서는 N-4.5에서 부분 감소하였다가 이후 N-12까지 증가하고 N-18에서 감소하였다(Fig. 1A). 질산태 질소 역시 차광조건에서는 N-9까지 증가하다가, N-12에서 급격하게 증가하였으며, N-18에서 감소하였으며, 비차광조건 역시 N-9에서 다소 감소하는 경향은 보였으나, 차광조건과 동일하게 N-12에서 급격히 증가하고 N-18에서 감소하였다(Fig. 1B). 차광조건이 비차광조건보다 암모늄태 질소는 1.3배, 질산태 질소는 1.9배 높았다. 차광조건의 경우, 차광망(95%)을 덮기때문에 공기순환이 정체되고, 비차광조건보다 상대습도 역시 8% 이상 높아 유기태 질소의 무기화가 촉진된 것으로 보여지며, N-12에서 높은 무기화를 보인 이유는, N-12는 타처리 유박보다 토양내에서 빠르게 무기화 및 질산화 과정을 거쳐 무기물(NH4+, NO3–)형태로 전환된 것으로 보인다. 질소는 동화작용을 통해 작물 생육에 반드시 필요하지만, 작물의 이용량 보다 과다할 경우, 작물의 질소 과잉 장애와 더불어, 환경적으로 악영향을 줄 수 있는데, 이중 질산태질소(NO3–)는 토양 내 이동성이 좋아 강우에 의해 용탈되기 쉽고 침수조건에서 탈질로 소실되기 쉬우며, 이는 지하수 및 지표수로 유입되어 부영양화를 발생시킬수 있다(Carpenter et al., 1998). 또한 대기로 손실될 경우 방출 가스 중 아산화질소(N2O)는 온실가스로 작용한다. 그 외에도 암모늄태 질소(NH4+)는 대기로 휘산 될 경우 미세먼지 원인물질로 작용할수 있다(Tsimpidi et al., 2007; Behera and Sharma, 2010).
질소함량 및 차광에 따른 찻잎 내 유리아미노산
아미노산은 차의 감칠맛을 나타내는 성분으로 카테킨의 쓴맛, 떫은맛과 더불어 차의 맛을 형성하는 중요한 요소이다(Kimura and Murata, 1971; Kakuda et al., 2000). 차 잎에 가장 많이 함유된 아미노산은 테아닌(theanine)이며 수용성으로서 좋은 품질의 녹차일수록 테아닌, 글루탐산(glutamic acid), 아르기닌(arginine) 등 다량의 아마이드(amide)가 함유되어있다(Cho et al., 2007). 차나무의 경우 질소 시비량이 증가할수록 유리아미노산 함량은 증가하고, 특히 theanine과 arginine이 크게 증가하였다(Ko et al., 2010). 이는 질소시비량이 많을수록 차나무 뿌리에서 질산염을 흡수하여 유기질소화합물로 동화하고(Marschner, 1995), 광호흡에 의해서 발생한 암모늄을 신속하게 아미노기 전달반응, 시트르산 회로, 암모늄 대사 등에 의해 다양한 아미노산을 다량 합성하기 때문이다(Lam et al., 1996). 또한 차광재배시 비차광 대비하여 높은 엽록소함량, 광합성 반응을 통해 생성된 광에너지가 광호흡작용에 필요한 에너지를 공급함에 따라 암모늄 대사를 통해 아미노산이 다량 합성되어 총아미노산함량 3.4배, 이중 theanine 3.7배, arginine 10.8배까지 증가하였다(Hwang et al., 2018). 총 유리아미노산 함량은 비차광 N-0에서 가장 낮은 199.7mg·100g-1으로 N-4.5에서 1.2배, N-9에서 2.1배, N-12에서 1.8배, N-18에서 3.1배 증가한 614.0mg·100g-1으로 조사되었다(Table 3). 차광 N-0의 총 유리아미노산은 401.7mg·100g-1으로 비차광 N-0 대비 2.0배 증가하였으며, N-12를 제외하면 질소 농도가 증가할수록 총 유리아미노산 함량이 비차광 N-0 대비 2.0–5.4배까지 증가하였다. 유리아미노산 중 암모니아의 경우, 차광 N-12에서 91.8, 차광 N-18에서 110.6, 비차광 N-18에서 99.3mg·100g-1으로 높게 조사되었다. 차광조건하에서 속효성이 N-12 유박의 경우, arginine, proline, theanine, glutamic acid, taurine 등 유용 유리아미노산이 N-9, N-18 보다 감소하는 패턴을 보인다.
Table 3.
Free amino acid content (mg·100 g-1) of tea leaves according to nitrogen content and shading treatment
| Tretment | NS | S | |||||||||
| 0.0% | 4.5% | 9.0% | 12.0% | 18.0% | 0.0% | 4.5% | 9.0% | 12.0% | 18.0% | ||
| Taurine | 0.11 bz | 0.18 a | 0.20 a | 0.12 b | 0.21 a | 0.14 a | 0.11 ab | 0.10 b | 0.11 a | 0.12 ab | |
| Phosphoethanolamine | 0.38 b | 0.43 a | 0.27 c | 0.17 d | 0.14 d | 0.40 b | 0.44 a | 0.22 d | 0.25 d | 0.33 c | |
| β-alanine | 0.46 a | 0.24 c | 0.41 b | 0.48 a | 0.41 b | 0.40 c | 0.50 b | 0.27 d | 0.48 b | 0.59 a | |
| β-Aminoisobutyric Acid | 0.43 b | 0.69 a | 0.21 d | 0.18 d | 0.26 c | 0.37 a | 0.12 c | 0.18 b | 0.15 bc | 0.19 b | |
| Methionine | 0.62 b | 0.81 a | 0.50 c | 0.64 b | 0.71 ab | 0.71 c | 0.48 d | 0.54 d | 0.96 b | 1.25 a | |
| Ornithine | 0.87 c | 0.52 c | 6.02 b | 0.87 c | 11.83 a | 0.48 d | 1.28 c | 4.02 b | 0.30 d | 9.53 a | |
| Glycine | 0.68 c | 0.78 c | 1.05 b | 1.18 b | 1.44 a | 1.24 a | 1.24 a | 1.17 a | 1.32 a | 1.45 a | |
| Valine | 1.38 c | 1.44 c | 1.46 c | 2.49 b | 3.83 a | 7.82 b | 3.10 e | 6.23 c | 5.15 d | 13.10 a | |
| L-Isoleucine | 1.35 ab | 1.30 b | 0.52 d | 1.50 a | 1.06 c | 4.55 a | 3.06 c | 2.24 d | 2.88 c | 3.61 b | |
| Cystine | 1.61 a | 1.63 a | 0.90 c | 1.47 b | 0.64 d | 2.02 a | 1.32 b | 1.27 b | 0.66 c | 0.82 bc | |
| L-Leucine | 1.74 b | 2.12 ab | 1.83 b | 1.79 b | 2.39 a | 2.91 c | 2.07 d | 3.89 b | 2.54 c | 4.43 a | |
| L-Tyrosine | 1.88 bc | 2.17 ab | 1.71 c | 1.59 c | 2.22 a | 8.15 a | 5.73 b | 5.27 bc | 5.09 c | 7.86 a | |
| Phenylalanine | 2.22 a | 1.59 b | 0.76 d | 1.24 c | 0.84 d | 5.34 a | 3.57 c | 2.63 d | 3.43 c | 4.61 b | |
| Histidine | 2.54 b | 1.81 c | 2.26 b | 2.28 b | 6.84 a | 6.86 b | 4.43 d | 6.60 c | 4.67 d | 11.10 a | |
| γ-Aminobutyric Acid | 2.94 d | 3.16 c | 5.04 b | 6.49 a | 5.06 b | 6.11 c | 6.70 b | 5.24 d | 4.82 e | 10.68 a | |
| Phospho-L-serine | 3.19 d | 3.39 c | 3.12 d | 3.74 bc | 4.45 a | 3.78 ab | 3.47 bc | 3.13 c | 3.90 a | 3.96 a | |
| Lysine | 3.26 d | 2.69 e | 4.55 b | 3.65 c | 6.68 a | 6.02 b | 5.44 c | 6.06 b | 4.32 d | 11.90 a | |
| Arginine | 3.29 e | 5.25 d | 40.33 b | 24.45 c | 76.69 a | 20.11 e | 41.56 c | 68.93 b | 29.73 d | 140.94 a | |
| Asparagine | 3.35 c | 2.23 d | 3.33 c | 8.14 b | 11.01 a | 27.08 d | 30.75 c | 62.55 b | 63.37 b | 171.62 a | |
| L-Alanine | 4.71 d | 5.60 cd | 10.32 a | 9.65 b | 10.60 a | 22.36 cd | 33.70 a | 24.24 cd | 21.06 d | 29.63 b | |
| Tryptophan | 5.89 a | 5.91 a | 1.17 c | 3.51 b | 0.51 d | 7.84 a | 5.71 b | 4.54 c | 5.63 b | 5.57 b | |
| L-Threonine | 6.16 b | 4.44 d | 5.19 c | 5.25 c | 6.78 a | 17.51 b | 14.91 c | 13.11 d | 14.79 c | 18.72 a | |
| Proline | 5.84 a | 3.44 d | 3.99 c | 5.06 b | 6.30 a | 6.71 d | 7.95 c | 10.62 b | 6.33 d | 20.84 a | |
| Theanine | 14.63 e | 37.02 d | 106.32 b | 84.34 c | 140.31 a | 60.31 e | 101.60 c | 130.10 b | 87.86 d | 276.87 a | |
| L-Glutamic acid | 112.83 e | 128.36 d | 186.02 b | 166.45 c | 213.44 a | 167.75 d | 190.12 c | 225.81 a | 151.99 e | 212.563 b | |
| Ammonia | 17.36 c | 14.35 c | 35.71 b | 28.85 b | 99.34 a | 14.72 d | 31.36 c | 33.97 c | 91.83 b | 110.63 a | |
| Total | 199.72 e | 231.55 d | 423.19 b | 365.58 c | 613.99 a | 401.69 e | 500.72 d | 622.93 b | 513.62 c | 1072.903 a | |
질소함량 및 차광에 따른 찻잎 내 카테킨, 카페인 함량
차의 대표적인 페놀성 화합물인 카테킨은 차의 향과 맛 등의 품질에 영향을 주며, 콜레스테롤 억제, 항산화 작용, 항암, 항균 작용을 한다(Higdon and Frei, 2003; Mohan et al., 2007). 본 연구에서 조사한 5종(EGCG, ECG, EC, EGC, C)의 개별 카테킨류 함량을 합하여 제시한 총 카테킨 함량은 차광 N-0에서 9.2%, 차광 N-4.5에서 9.17%로 가장 높았으며, 비차광 N-12에게 6.18%로 가장 낮았다(Table 4). 이러한 카테킨 중 가장 많은 비율을 차지하고 있는 EGCG(epigallocatechin gallate)는 차광 N-4.5에서 4.05%로 가장 높았으며, 차광 –9.0~18.0까지 유사한 수준을 보였고, 비차광처리구에서는 N-18에서 3.07%로 가장 높은 경향을 보였으며, 차광과 비차광으로 구분할 때 차광처리에서, 질소함량별 처리구에서는 N-4.5에서 높은 EGCG 함량을 보였다. 일반적으로 차광조건에서 증가하는 테아닌은 gallate형 카테킨인 EGCG, ECG의 발현을 촉진하는데(Havaux et al., 1996; Goto et al., 1996), 이로 인해 차광처리구가 비차광처리보다 EGCG, ECG 함량이 1.3배, 1.5배 증가하였다. 기관지 및 혈관 확대, 이뇨작용, 생리적 스트레스 저하 등의 기능 및 차의 쓴맛에 관여하는 카페인은 차광 N-0에서 0.233%로 가장 높았으며, 차광 N-9, 12, 18, 비차광 N-0, 4.5에서 유사한 경향을 보였다. 전체적으로 차광 조건에서 과발현되는 테아닌에 직접적인 영향을 받아 차광조건에서 비차광 대비하여 1.23배 증가하였으며, 질소 시비시 오히려 감소하였다.
Table 4.
Catechin and caffeine content (%) of tea leaves according to nitrogen content and shading treatment
| Tretment | EGCz | C | EC | EGCG | ECG | Total catechin | Caffeine | |
| Shading | N content | |||||||
| NS | 0% | 2.680 ey | 0.018 ab | 0.647 f | 2.682 fg | 0.707 b | 6.733 e | 0.132 b |
| 4.5% | 2.657 e | 0.017 ab | 0.673 e | 2.813 ef | 0.447 e | 6.607 e | 0.133 b | |
| 9.0% | 2.648 e | 0.017 ab | 0.682 e | 2.593 g | 0.713 b | 6.653 e | 0.102 c | |
| 12.0% | 2.313 g | 0.015 ab | 0.682 e | 2.773 ef | 0.400 e | 6.183 f | 0.100 c | |
| 18.0% | 2.513 f | 0.013 ab | 0.627 g | 3.070 d | 0.455 e | 6.678 e | 0.073 d | |
| S | 0% | 3.462 a | 0.020 ab | 0.880 b | 3.463 c | 1.390 a | 9.215 a | 0.233 a |
| 4.5% | 3.503 a | 0.020 ab | 0.905 a | 4.050 a | 0.690 bc | 9.168 a | 0.063 d | |
| 9.0% | 3.178 b | 0.012 b | 0.780 c | 3.698 b | 0.598 cd | 8.267 b | 0.128 b | |
| 12.0% | 2.842 d | 0.017 ab | 0.770 c | 3.592 b | 0.572 d | 7.792 d | 0.112 b | |
| 18.0% | 2.938 c | 0.017 ab | 0.747 d | 3.603 b | 0.690 bc | 7.995 c | 0.128 b | |
결과적으로 말차 생산을 위한 차나무 차광 재배시 차나무 생육, 토양 유기태질소 변화, 찻잎의 유리아미노산 함량 등을 고려할 때, 차광 직전 과도한 질소 시비는 오히려 찻잎의 품질 저하의 우려가 크고, 2차적으로 차광망 내부의 과습, 고온 등의 기후조건과 더불어 세포조직의 연질화로 인한 병해충 발생에 취약할 수 있기 때문에 비교적 질소질 함량이 낮은 4.5% 질소를 시비하는 것이 바람직하다.
