

Introduction
The production value of Kimchi cabbage (Brassica campestris spp. pekinensis) in Korea in 2015 was approximately 663 billion won, which was 7.3% of the total vegetable production (MAF statistics, 2015). The cultivation area and yield of Kimchi cabbage represented 26,179 ha and 7,868 kg/10a, respectively. The major growing season for Kimchi cabbage is in the autumn between late September and November, which accounted for 70% of the total yield, followed by the spring (14%) and winter (9%), and cultivation in alpine areas (7%), respectively.
Low temperature is one of the most important abiotic stresses that affect plant growth and development (Thakur et al., 2010; Zhang et al., 2013). The photosynthesis with treated cold stress of warm-climate plant is the most significantly affected biological process (Allen and Ort, 2001; Goh et al., 2012). Cold stress has deleterious effects on the capacity and efficiency of photosynthesis that are attributed to altered gas exchange, pigment content and composition, and chloroplast development, as well as the loss of chlorophyll in source leaves (Farooq et al., 2009). Low temperatures also induce the overproduction of reactive oxygen species (ROS) that can promote oxidative stress in lipids, proteins, and nucleic acids (Mittler, 2002). ROS include singlet oxygen (1O2), superoxide anion (O2-), hydroxyl radical (·OH), and hydrogen peroxide (H2O2), which are major effectors of oxidative cellular metabolism. Additionally, H2O2 accumulation may function in various biotic or abiotic stresses (Kuźniak and Urbanek, 2000).
The activity of antioxidant enzymes such as superoxide dismutase (SOD), ascorbate peroxidase, catalase (CAT), and peroxidase (POD), and antioxidant metabolites like ascorbate, tocopherol, and glutathione can play crucial roles in controlling and preventing stress-induced accumulation of ROS (Foyer and Noctor, 2005; Suzuki and Mittler, 2006; Turhan et al., 2008). SOD is a major O2- scavenger and its enzymatic action results in the reduction of H2O2 and O2-. H2O2 is scavenged by several peroxidases and CAT, which dismutates H2O2 into H2O and O2 (Mckersie and Leshem, 1994). Since this reaction requires the simultaneous access of two H2O2 molecules in the active site, CAT has extremely high maximum catalytic rates (Willekens et al., 1995). POD, which has less specificity to electron donor substrates, decomposes H2O2 by the oxidation of co-substrates such as phenolic compounds (Mehlhorn et al., 1996). These ROS-scavenging enzymes improve stress tolerance through diminishing the harmful effects of ROS. Indeed, cells with higher levels of antioxidant capacity had greater resistance to different types of environmental stresses (Dat et al., 1998; Thomas et al., 1999). Glutamic acid can play a substantial role as the precursor of proline biosynthesis, which is a cyclic amino acid that accumulates following a variety of environmental stresses, including drought, extreme temperatures, salinity, and heavy metal exposure (Munns, 2005; Sharma and Dietz, 2006). However, characteristic changes in photosynthesis and antioxidant systems that are induced by glutamic acid in low air temperatures are not well defined. The present study aimed to ascertain the physiological responses of Kimchi cabbage cultivars to exogenously applied glutamic acid when exposed to low temperature stress, and to clarify whether glutamic acid could contribute to alleviating coldinduced physiological damage.
Materials and Methods
The Kimchi cabbage seeds ‘Chunkwang’(Sakada Seed Co., Japan) were sown on June 9, 2016 in a plug tray with a single cell volume of 35 mL, filled with commercial soil (Bio Media, Heungnong Seed Co., Korea). The transplanting was carried out in a glasshouse at the National Institute Horticultural and Herbal Science, Wanju, Korea (35o16’N, 127o02’E, 32 m above sea level). The plants were transplanted 23 days after sowing into the 4,735 × 4,625 × 3,860 mm (L×W×H, outside) extreme weather growth-chambers (EWGC; Modified CEEWS model, EGC, Chagrin Falls, OH, USA). Twenty-eight days after transplanting (DAT) and when the plants had about 35 leaves, the Kimchi cabbages were introduced to a low (10°C) air temperature for 7 d. Plants were then treated once with 200 mL/plant of pure water, 0.3% urea, or 10 μmol·mol-1 glutamic acid solutions.
After the low air temperature treatment, the internal leaves of the Kimchi cabbages at 0, 3, and 8 d were collected. After the glutamic acid treatments, the leaves were harvested another six times, until the 28th day. After the foliar treatments, non-shaded or 30%-shaded conditions were applied using mesh nets. The fresh weights of the shoot, head, and root were measured at 61 DAT for each cabbage. The yields were calculated as the planting density (2,850 plants/10 a) × fresh weight of the cabbage head × percentage of head formation (100% = 1.0, 0% = 0.0). The CO2 assimilation rate (An) response to the intercellular CO2 concentration (Ci) (An-Ci) curve and An responses to the light curve of the 0, 1, 3, and 5 d after cold treatment were measured. Kimchi cabbage external leaves were measured using a portable photosynthesis system (Li 6400, Li-Cor Co., Inc., Lincoln, NE, USA). For the An-Ci curve, gas exchange responses to various CO2 concentrations ranging from 50 to 1,100 μmol·mol-1 were measured at 500 μmol·m-2·s-1 photosynthetic photon flux (PPF). Biochemical models (Farquhar et al., 1980) were used to determine net photosynthetic rate, which is limited by the rates of CO2 carboxylation by rubisco (ribulose 1, 5-bisphophate; RuBP) regeneration and triose phosphate utilization (TPU). These limiting photosynthetic parameters of the Kimchi cabbage leaves were calculated using an Excel macro program developed by Sharkey et al. (2007). In addition, we measured photosynthesis characteristics a total of eight times at 10, 13, 16, 19, 21, 25, 28, and 31 d after cold treatments, respectively. The photosynthesis characteristics were represented as an average of the eight measurements. The portable Li-Cor gas exchange system maintained flow rate and photosynthetic photon flux at 500 μmol·s-2 and 500 μmol·m-2·s-1, respectively. Leaf temperature was also set at 25°C and relative humidity (RH) at 40-60%. The leaf area in the leaf chamber was 600 mm2. All the measurements were replicated three times.
The plant tissue of external leaves in the replicated trials were harvested and then directly frozen in liquid nitrogen. Fresh leaves (50 mg) were ground to a fine powder in a mortar using liquid nitrogen and then soaked in a 20 mM potassium phosphate buffer (pH 6.5) on ice. The extracts were clarified by centrifugation at 13,500 g for 15 min at 4°C, and then stored at –80°C until analysis.
The total proteins from the Kimchi cabbage leaves were extracted according to the method of Bradford (1976), using bovine serum albumin (BSA) as a standard. The superoxide dismutase (SOD) activity, which catalyzes the conversion of superoxide to H2O2, was determined using an SOD assay kit-WST [2-(4-iodophenyl)-3-(4-(nitrophenyl)-5-(12.4-disulfophenyl)- 2H-tetrazolium, mono-sodium salt, Sigma #19160] according to the manufacturer’s instructions. The catalase (CAT) activity was measured by using an Amplex Red Catalase Assay Kit (Molecular Probes, Eugene, OR, USA). The CAT activity was determined by measuring the absorbance at 560 nm using a microplate reader (Eon-C, Biotek, Winooski, VT, USA). The H2O2 concentration and peroxidase (POD) activity were measured from sample supernatants using an Amplex Red hydrogen peroxide/peroxidase assay kit according to the manufacturer's instructions (Molecular Probes). Initially, reaction mixtures containing 50 μL peroxidase-containing samples (diluted 2x) and 50 μL of 100 μM Amplex Red reagent containing 2 mM H2O2 were incubated at room temperature for 30 min in the dark. The H2O2 concentration and POD activity were determined by measuring the absorbance at 560 nm using a microplate reader (Eon-C, Biotek, Winooski, VT, USA).
A total of 30 plants were placed in each EWGC treatment condition and five biological replicates of the six treatments were arranged using a completely randomized design. An analysis of variance (ANOVA) was performed with SAS (SAS 9.2, SAS Institute Inc., Cary, NC, USA). The mean separation for the growth and photosynthetic parameters was conducted using Duncan’s multiple range tests (p <0.05).
Results
Of the treatment conditions, the maximum cabbage fresh weight (1,700 g/plant) was measured in the glutamic acid treatment and the minimum fresh weight (1,193 g/plant) in the urea with shading treatment. The head weight and yield of cabbages treated with glutamic acid were 854.6 g/plant and 4,845 kg/10 a ares, respectively, which were the highest among all the treatments tested. The smallest yield (3,400 kg/10 ares) was observed in cabbages treated the urea and shading (Table 1). Sixty-one days after transplanting (the 28th day after foliar treatments), the greatest leaf length was obtained in the shaded plants, while the plants treated with urea in shaded conditions produced plants with the smallest leaf length (data not shown). These results were similar to previously reported results, in which growth and yield were reduced, while the survival in cucumbers (Nam et al., 2006) and tomatoes (King et al., 1982) improved with ABA treatment under low air temperatures.
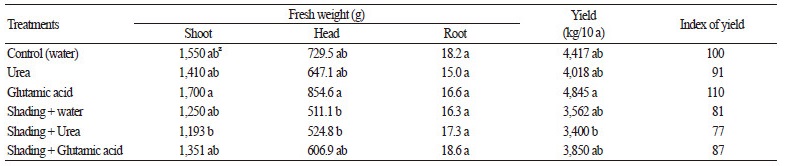

Table 3. Net photosynthesis of Kimchi cabbage treated with foliar sprays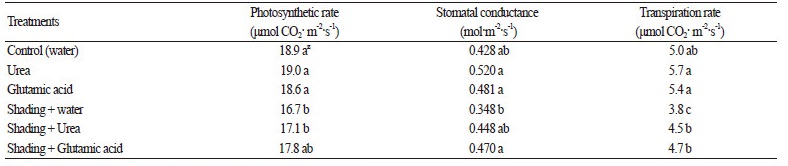 |
z Mean separation within columns by Duncan’s multiple range test at p < 0.05. |
The highest carboxylation and electronic transport rates were 186 μmol·m-2·s-1 and 133 μmol·m-2·s-1, respectively, at one day after cold treatment (Table 2). TPU was significantly highest before low air temperature treatment, while mitochondrial respiration was not available to calculate after cold treatment (Table 2). In addition, the net photosynthetic rate and stomatal conductance were the highest with the glutamine and urea spray treatments; however, they decreased in the plants gown in the shading treatment (Table 3). Low temperature stress of Kimchi cabbages adversely affected the net photosynthetic rate and stomatal conductance; however, there was an increase in the carboxylation rate and photosynthetic electron transport system efficiency. The net photosynthetic rate in Kimchi cabbage leaves that were treated with low air temperatures was 13 μmol CO2·m-2·s-1, which was 28% lower than that measured before the low air temperature treatment (data not shown). These results are similar previous studies that showed that photosynthesis is the most significantly affected by cold temperature injury (Allen and Ort, 2001; Goh et al., 2012; Lee et al., 2016; Park et al., 2014). Cold stress decreases the capacity and efficiency of photosynthesis by changing gas exchange (Farooq et al., 2009).
The damage to Kimchi cabbages was decreased with glutamic acid spray treatment; however, it was not decreased with glutamic acid treatment in the shade (Fig. 1). The leaves of Kimchi cabbage were observed through an optical microscope and showed normal epidermal tissue (i.e. non-shrinked and non-breaked cell) after the glutamic acid and control (water) treatments. The glutamic acid in combination with shading treatment produced abnormalities at the epidermal tissue, palisade, and spongy parenchyma (Fig. 1).
The SOD activity was slightly increased under cold treatment, while the foliar sprays and shading treatments did not have a significant effect on SOD activity (Fig. 2A). In the 3 d and 8 d after low air temperature treatment, the POD activity increased 2-fold with cold treatment, while the activity was lowest 15 d after the glutamic acid treatment (Fig. 2B). The CAT activity in the early stages (3 d) more than tripled after low air temperature treatment and then reverted to the base line quickly after treatment with glutamic acid (Fig. 2C). Notably, the H2O2 content was the lowest after glutamic acid treatment (Fig. 2D). Low air temperature caused an increase in the antioxidant enzyme activity in Chinese cabbage seedlings and the application of exogenous nitric oxide further enhanced the antioxidant enzyme activities (Fan et al., 2014). Our results did not show a further increase in any other antioxidant enzyme after glutamic acid treatment, but the antioxidant enzyme activities returned to their base line after application of glutamic acid more quickly compared to the other treatments tested. However, it is still unclear whether the glutamic acid directly or indirectly affected the responses to cold stress in the plants.
Proline is one of the most important organic osmolytes in plant cells and is rapidly accumulated in plants after cold stress (Liu et al., 2009; Liu et al., 2013). Glutamic acid can act as the precursor of proline (Munns, 2005; Sharma and Dietz, 2006). This may owe due to the fact that glutamic acid activates proline biosynthetic enzymes (Fariduddin et al., 2009, which was considered in conjunction with the increased CAT and POD activities that can act to improve the rate of recovery in the early stages of cold stress. Exogenous glutamic acid foliar treatments could alleviate cold-induced physiological damage by combining photosynthetic regulation and increased antioxidant capacity during cold stress. In addition, application of glutamic acid might be feasible chemical treatment methods for maintain the growth and yield of Kimchi cabbage under abiotic stresses.
Fig. 2.
Effects of foliar treatments on antioxidant enzyme activity in terms of SOD (A), POD (B), CAT (C) activity and H2O2 content (D) in leaves of Kimchi cabbage grown at 10°C air temperature. Bars represent standard errors of three replications.
