

서 언
재료 및 방법
실험 재료 및 장소
생장조절제 및 빛 처리
생장조절제 및 빛 처리에 따른 가지 세력 및 액아 크기 측정
생장조절제 및 빛 처리에 따른 액아의 화원기형성률 및 눈 괴사율 측정
통계 분석
결과 및 고찰
환경휴면기의 생장조절제 및 빛 처리에 따른 액아의 특성
생육기의 생장조절제 및 빛 처리에 따른 가지와 액아의 특성
서 언
포도의 액아는 하나의 주아와 두개 이상의 부아로 이루어져 있으며, 잎으로 분화되는 엽원기와 꽃으로 분화되는 화원기를 함께 가지고 있다. 액아 내부에는 품종에 따라 10-12개의 엽원기와 1-3개의 화원기가 동시에 형성된다. 특히 화원기는 빛, 온도, 수분 등의 환경요인(Deloire, 2009; Vasconcelos et al., 2009)이나 gibberellin과 cytokinin 등과 같은 호르몬(Vasconcelos et al., 2009)에 의해 분화된 후 겨울 휴면기 동안 과종 혹은 품종에 따라 다양한 휴면심도의 변화 과정을 거친 후, 이듬해 봄에 발아하기 전까지 발달을 지속한다(Williams, 2000; Park and Park, 2020). 포도의 잠재적인 생산량은 나무 당 송이가 달리는 가지의 수에 크게 영향을 받는다(Hopping, 1977). 더 나아가 가지의 수가 충분한 경우에도 화수가 없는 빈 가지가 발생하는 경우가 있어 송이가 되는 액아의 화원기 형성이 충실해야 생산량이 증가한다.
액아의 형성 및 발달은 다양한 요인에 영향을 받는다. ‘Thompson Seedless’, ‘Flame Seedless’, ‘Chardonnay’, ‘Cabernet Sauvignon’ 등의 포도 품종의 경우, 빛에 많이 노출될수록 화원기 형성이 증가한다고 보고되었다(Sánchez and Dokoozlian, 2005). 반면에 액아에 탄수화물의 공급이 부족할 경우, 원기(primordia)의 괴사가 일어난다(Lebon et al., 2008). 수분 또한 화원기의 수와 크기에 영향을 미치면서 수분 스트레스가 지속된다면 액아의 생산력이 저하된다. 식물 생장조절물질인 gibberellic acid(GA)는 처리 시기에 따라 액아의 화원기 형성을 억제하기도 하며, cytokinin 처리는 화원기 형성을 유도한다고 보고되었다(Li-Mallet et al., 2016). 담배의 경우 잎눈보다 꽃눈에서 polyamine류인 spermidine(SPD)이 많이 검출되었으며, SPD을 처리할 경우 꽃눈의 분화가 야기되었다(Kaur-Sawhney et al., 1988).
‘샤인머스캣’ 포도는 1988년 일본에서 ‘Akitsu-21’(Vitis labruscana)과 ‘Hakunan’(Vitis vinifera)을 교배하여 육성한 이배체의 청포도 품종으로 품질이 매우 좋고 무핵재배가 가능하다. 만부병, 노균병, 흰가루병에는 저항성이 있지만 탄저병에는 감수성을 가진다(Yamada et al., 2008). 많은 장점에도 불구하고 ‘샤인머스캣’ 포도는 신초의 액아에서 꽃송이가 나오지 않는 빈가지 발생이 빈번하여 생산량을 감소시키는 문제점이 있으나, 현재까지 ‘샤인머스캣’ 포도의 빈가지 발생의 원인과 경감 방법에 대한 연구는 거의 없는 실정이다.
따라서 본 연구에서는 ‘샤인머스캣’ 포도의 빈가지 발생을 경감시키고자 식물 생장조절제인 6-benzylaminopurine(BAP)과 spermidine(SPD) 그리고 환경요인인 빛 처리를 통해 액아의 발달을 보았다.
재료 및 방법
실험 재료 및 장소
본 연구는 2019년과 2020년에 경기도 안성시와 충청남도 천안시 소재의 개인 농가에서 ‘샤인머스캣’ 포도를 대상으로 실험을 진행하였다.
생장조절제 및 빛 처리
2019년의 실험은 환경휴면기의 처리와 생육기의 처리로 나누어 진행하였다. 환경휴면기인 3월 중순에 4년생 ‘샤인머스캣’포도에서 처리구당 발아 전인 1년생 가지 30개를 채취한 후 4번 액아부터 8번 액아까지를 이용하여 70% 에탄올로 소독하고, 플로랄폼에 꽂아 7일간 온도 25°C인 환경에서 수삽하였다. 생장조절제로는 BAP(6-benzylaminopurone, Sigma-Aldrich, USA) 200ppm(Nambiar et al., 2012)과 SPD(Spermidine, Sigma-Aldrich, USA) 140ppm(Khezri et al., 2010)을 각각 수삽 전 1회 침지 처리하였으며, 생장조절제 처리와 별도로 환경요인의 영향을 보기 위하여 7일의 수삽 기간 동안 매일 18시간의 장일처리를 하여 수삽 전·후의 액아의 크기 변화와 화원기형성률을 조사하였다. 많은 포도 품종에서 빛에 많이 노출될수록 화원기 형성이 증가한다는 Sánchez and Dokoozlian(2005)의 보고와 모든 포도 품종에서 단일조건이 과실의 품질과 수량을 현저히 저하시킨다(Yim, 2020)의 보고를 참고하여 단일처리는 실험에서 배제하였다. 실내에서의 수삽 실험을 통해 액아의 화원기형성률을 조사하는 한편, 실제 포장에서의 빈가지 발생률을 확인하기 위하여 실내 실험과 동시에 동일한 처리를 포장에도 실시하였다.
생육기 처리는 포장에서만 진행하였으며, 가지 발달, 액아 발달 그리고 화원기형성률을 보기 위해 신초의 마디가 10마디 이상 형성된 6월에 3년생 ‘샤인머스캣’포도 12주를 대상으로 처리당 3반복으로 환경휴면기와 동일하게 BAP 200ppm, SPD 140ppm 및 18시간의 장일처리를 하였다. BAP 처리와 SPD 처리는 6월에 각 1회 스프레이를 이용하여 가지 전체에 처리하였으며, 장일처리구에는 전등을 설치한 후, 빛이 다른 나무들에 영향을 미치지 못하도록 처리구 주변에 폴리에스테르와 면 혼방 재질로 된 검은색 암막을 설치하였다. 장일처리구 또한 암막 처리로 인한 주간의 광투과와 야간의 온도 변화 등에 영향이 최소화되도록 상부는 열린 채로 두고 장일처리구로부터 3m 이상 충분히 거리를 두고 암막을 설치하였다. 18시간의 장일을 정확히 처리하기 위해 일몰 후 22시까지 빛을 주었고, 다음날 04시부터 일출 전까지 빛 처리를 하였다. 즉, 22시부터 다음날 04시까지를 제외한 모든 시간에 포도나무가 자연광 또는 전등에 의해 빛에 노출되도록 하였으며, 6월부터 수확기까지 지속적으로 처리하였다. 처리구당 12개의 신초를 생장조절제 및 빛 처리 전·후에 생육기간별로 채취하였으며 이듬해인 2020년 3월에 마지막으로 채취하여 분석하였다. 1번 액아부터 10번 액아까지를 실험에 사용하였으며, 가지의 세력, 액아의 크기, 화원기형성률과 눈 괴사율을 조사하였다. 단 2020년 3월에 채취한 가지의 경우, 현재 농가에서 주로 행해지는 단초전정을 하였기에 2번 액아까지 조사하였다.
2020년 3월(환경휴면기)에는 2019년 환경휴면기의 처리 실험의 결과를 토대로 화원기형성률이 가장 높았던 BAP를 선택하여 BAP 농도에 따른 액아 변화와 화원기형성률을 조사하였다. 수삽은 5년생 ‘샤인머스캣’포도에서 처리구당 가지 60개를 2020년 3월 5일부터 10일 간격으로 총 3번 채취하였고, 가지당 4번 액아부터 8번 액아까지 사용하여 2019년과 동일한 방법으로 진행하였다. BAP는 시료 채취시기마다 100ppm, 200ppm, 400ppm을 각각 처리하였다.
생장조절제 및 빛 처리에 따른 가지 세력 및 액아 크기 측정
액아의 종경과 횡경은 Vernier Calipers(CD-15CPX, Mitutoyo corp., Japan)를 이용하여 측정하였다.
생육기인 2019년 6월부터 채취한 가지의 생장조절제 및 빛 처리에 따른 생장 정도를 알아보기 위해 가지의 1번 마디부터 10번 마디 사이의 굵기와 1번 마디부터 10번 마디까지의 길이를 측정하였고, 1번 액아부터 10번 액아의 종경과 횡경의 길이를 측정하였다. 2020년 3월에 채취한 가지는 1번, 2번 액아의 종경과 횡경의 길이만 측정하였다. 가지 길이는 줄자를 이용하여 측정하였고, 가지 굵기와 액아의 종경, 횡경은 Vernier Calipers(CD-15CPX, Mitutoyo corp., Japan)를 이용하여 측정하였다.
생장조절제 및 빛 처리에 따른 액아의 화원기형성률 및 눈 괴사율 측정
액아를 예리한 칼을 이용하여 종으로 절단한 뒤, 실체현미경(SZX10, Olympus, Japan)을 이용하여 화원기형성률과 눈 괴사율을 조사하였다.
통계 분석
수집된 데이터의 통계분석은 PASW statistic 18(SPSS inc., USA) 프로그램을 이용하여 Duncan 다중검정(p < 0.05)으로 분석하였다.
결과 및 고찰
환경휴면기의 생장조절제 및 빛 처리에 따른 액아의 특성
발아 전 환경휴면기인 2019년 3월의 생장조절제 및 빛 처리가 액아의 크기와 화원기 형성에 어떤 영향을 미치는지 알아보기 위해 처리 후 7일간 수삽한 후 액아의 크기 변화와 화원기형성률을 조사하였다.
모든 처리구에서 액아 크기의 변화율이 양의 값을 가지는 것으로 보아, 액아의 크기 생장이 일어난 것을 알 수 있었으나 처리구간 통계적 유의성이 인정되지 않아 처리에 따른 변화보다는 환경휴면이 타파되어 액아의 생장이 진행된 것으로 생각되었다. 화원기형성률의 경우, BAP 처리를 할 경우 다른 처리구에 비해 화원기형성률이 높은 것으로 조사되었다. 반면에 18시간의 장일 처리에서는 화원기 형성이 억제된 것을 확인하였다. SPD 처리 시, 화원기형성률이 수치적으로 증가하였지만, 통계적 유의성은 없었다(Table 1).
Table 1.
The bud size change rate and inflorescence primordium formation from bud positions 4 to 8 by growth regulators and light treatments in March before germination of ‘Shine Muscat’ grapevine
| Treatment | Rate of Bud Size Changez |
Inflorescence primordium (%) | |
| Rate of change in bud length (%) | Rate of change in bud diameter (%) | ||
| Control | 2.56 ay | 1.41 a | 50.44 ab |
| BAP | 2.33 a | 0.36 a | 60.89 a |
| SPD | 2.01 a | 0.19 a | 56.00 ab |
| Long-Day | 3.04 a | 1.04 a | 44.00 b |
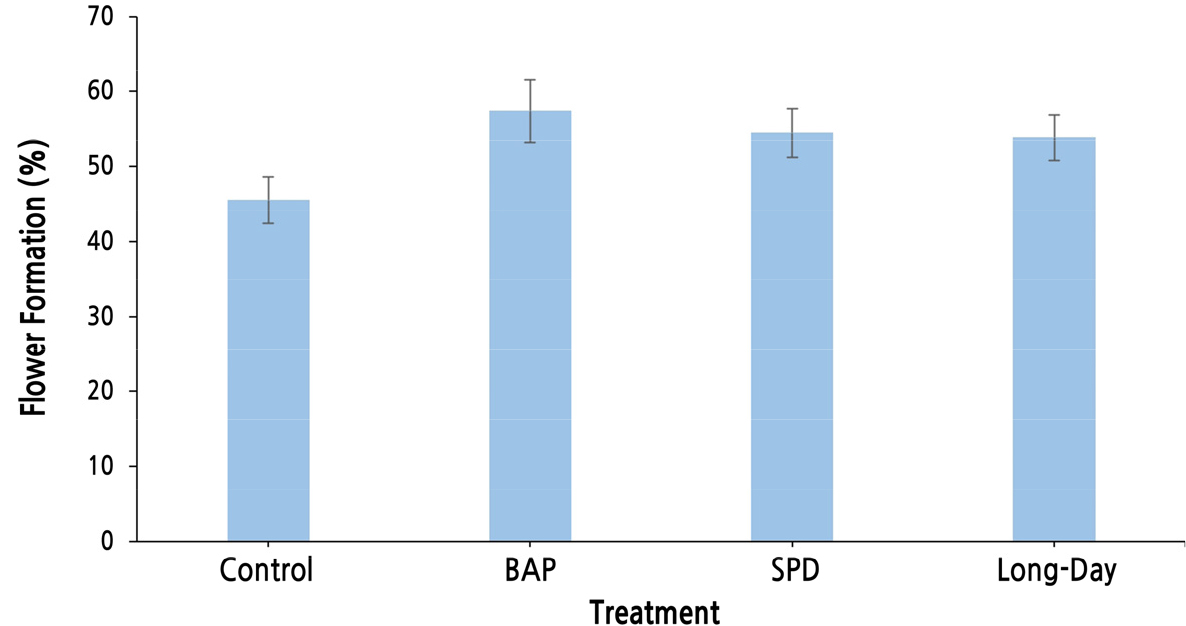
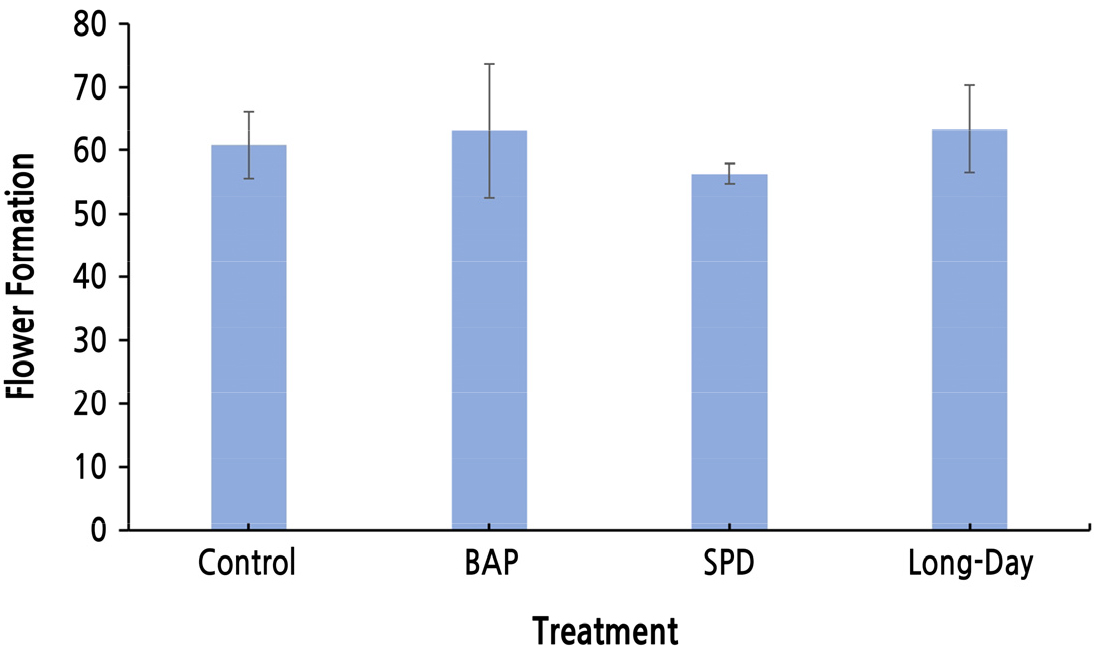
환경휴면기의 생장조절제 처리와 빛 처리는 액아의 크기 생장에는 영향을 미치지 않았으나, 반면에 화원기 형성의 경우, 장일 처리시 억제되고 BAP 처리시 유의하게 증가하는 것을 알 수 있었다. 그러나 이는 7일이라는 단시간 내의 화원기 형성을 본 것이기 때문에, 실제 빈가지가 발생하는 정도를 확인하기 위해 실험포장에서 동일한 실험을 진행하였다. 발아 후 꽃송이가 형성된 신초의 비율(전체 신초 수에 대해 꽃송이가 형성된 신초의 비율)은 무처리, BAP, SPD 및 장일 처리 시 각각 45.45%, 57.34%, 54.39%, 53.79%로 확인되었다(Fig. 1). 이를 통해 환경휴면기의 BAP 처리는 화원기 형성 속도와 형성률을 증가시키며, 장일 처리의 경우 화원기 형성 속도는 느려지지만 형성이 억제되지는 않는 것으로 생각된다.
BAP 200ppm이 화원기형성률을 증가시킨다는 결과를 토대로 2020년 3월에 BAP 농도에 따른 액아의 크기 변화와 화원기형성률을 보았다. 모든 처리구에서 액아의 크기 변화율이 양의 값을 보였지만 통계적 유의성은 나타나지 않은 것을 보아 처리 농도는 액아의 크기 생장에 영향을 미치지 않는 것으로 조사되었다. 화원기형성률의 경우, 통계적 유의성은 없었지만 BAP 200ppm에서 56.67%로 가장 높은 화원기형성률을 보였고 고농도인 BAP 400ppm에서 45.81%로 가장 낮은 형성률을 보였다(Table 2). BAP 농도뿐만 아니라 처리시기도 화원기 형성에 영향을 미칠 수 있다는 가정하에 처리 시기와 BAP 농도에 따른 화원기형성률을 비교해 보았다. 환경 휴면 타파 직후인 3월 5일에 처리한 BAP 200ppm이 63.33%로 가장 높은 화원기형성률을 보였으며, 그에 반해 역시 3월 5일에 BAP 400ppm을 처리한 처리구에서 화원기형성률이 40.00%로 가장 낮았다. 처리 시기가 늦어질수록 농도에 상관없이 비슷한 수준의 화원기 형성을 보인다는 것을 알 수 있었다(Table 3). 2년간의 결과를 통해 BAP 200ppm이 화원기형성률을 증가시키며, 특히 내재휴면 타파 후 빨리 처리하는 것이 가장 효과적이라고 생각된다.
Table 2.
The bud size change rate and inflorescence primordium formation of ‘Shine Muscat’ grapevine from bud positions 4 to 8 after 7 days of BAP treatment during ecodormancy in March
| Treatment | Rate of Bud Size Changez | Inflorescence primordium (%) | |
| Rate of change in bud length (%) | Rate of change in bud diameter (%) | ||
| BAP 100ppm | 0.58 ay | 0.41 a | 54.44 a |
| BAP 200ppm | 1.55 a | 0.77 a | 56.67 a |
| BAP 400ppm | 0.57 a | 0.98 a | 45.81 a |
Table 3.
Inflorescence primordium formation of ‘Shine Muscat’ grapevine according to the timing of treatment and BAP concentration
| Treatment | Inflorescence primordium (%) | |
| Dates | Concentrations (ppm) | |
| 5 March | 100 | 51.67 abz |
| 200 | 63.33 a | |
| 400 | 40.00 b | |
| 15 March | 100 | 56.67 ab |
| 200 | 51.67 ab | |
| 400 | 51.67 ab | |
| 25 March | 100 | 55.00 ab |
| 200 | 55.00 ab | |
| 400 | 45.76 ab | |
생육기의 생장조절제 및 빛 처리에 따른 가지와 액아의 특성
발아 후 생육기에 생장조절제 및 빛 처리를 할 경우, 가지와 액아의 생장과 액아 속 화원기 형성에 어떤 영향을 미치는지 알아보기 위한 실험을 진행하였다.
가지의 굵기는 1번 마디부터 10번 마디까지 마디 사이의 굵기를 평균내어 비교하였다. 7월, 8월, 10월 3개월간 월별로 가지의 굵기를 비교한 결과 모든 시기에서 처리구간 통계적 유의성이 인정되지 않아, 생장조절제와 빛 처리는 가지의 굵기 생장에는 영향을 미치지 않는 것으로 조사되었다(Table 4).
Table 4.
Comparison of shoot diametersz in ‘Shine Muscat’ grapevine according to growth regulators and light treatments in the growing period
| Treatment | Shoot diameter in July (mm) | Shoot diameter in August (mm) | Shoot diameter in October (mm) |
| Control | 7.49 ay | 9.31 a | 9.68 a |
| BAP | 8.60 a | 8.19 a | 10.89 a |
| SPD | 8.81 a | 9.20 a | 10.63 a |
| Long-Day | 7.82 a | 8.35 a | 10.82 a |
가지의 길이는 1번 마디부터 10번 마디까지의 총 길이를 측정하였다. 7월에는 처리구 간 통계적 유의성이 나타나지 않았으나, 8월부터 SPD을 처리한 가지의 길이 생장이 촉진된 것을 확인하였다(Table 5). SPD을 비롯한 polyamines이 세포분열에 관여(Carbonell and Blázquez, 2009)한다고 알려져 있어, SPD의 처리는 신초의 생장점의 세포분열을 촉진하여 길이 생장이 다른 처리구에 비해 높았던 것으로 생각되었다.
Table 5.
Comparison of shoot lengthsz of ‘Shine Muscat’ grapevine according to growth regulators and light treatments in the growing period
| Treatment | Shoot length in July (cm) | Shoot length in August (cm) | Shoot length in October (cm) |
| Control | 72.13 ay | 73.27 b | 69.45 b |
| BAP | 82.97 a | 71.81 b | 84.28 ab |
| SPD | 79.92 a | 91.33 a | 89.57 a |
| Long-Day | 73.07 a | 75.16 b | 80.63 ab |
SPD 처리는 또한 생육기의 액아 크기를 가장 빠르게 증가시켰다. 반면에, SPD와 마찬가지로 세포분열을 촉진시키는 것으로 알려진 BAP 처리는 생육기의 액아 생장을 늦추는 것으로 조사되었다. 따라서 BAP와 SPD 모두 세포분열에 관여(Carbonell and Blázquez, 2009; Ferrer et al., 2017)하나 작용기작이 다를 가능성이 확인되었으며, 이에 대한 더 깊이 있는 연구가 필요할 것으로 생각되었다. 장일 처리의 경우, 생육 후기인 8월 이후부터 액아의 크기가 증가하였으며 장일 처리의 영향은 일장이 짧아지는 후기로 갈수록 크게 나타나는 것으로 추정되었다(Table 6).
Table 6.
Comparison of bud lengths and diameters of 'Shin Muscat' grapevine according to growth regulators and light treatments in the growing period
| Treatment | Junez | July | August | October | |||||||
| Bud length (mm) | Bud diameter (mm) | Bud length (mm) | Bud diameter (mm) | Bud length (mm) | Bud diameter (mm) | Bud length (mm) | Bud diameter (mm) | ||||
| Control | 3.26 | 2.89 | 3.96 by | 3.69 bc | 4.71 a | 4.22 a | 4.62 b | 3.88 a | |||
| BAP | 3.26 | 2.89 | 4.20 a | 3.86 b | 4.11 b | 3.80 b | 4.75 b | 3.96 a | |||
| SPD | 3.26 | 2.89 | 4.41 a | 4.10 a | 4.60 a | 4.05 a | 4.69 b | 3.85 a | |||
| Long-Day | 3.26 | 2.89 | 3.87 b | 3.56 c | 4.49 a | 4.18 a | 5.05 a | 4.04 a | |||
눈 괴사율은 모든 조사 시기에서 처리에 따른 통계적 유의성이 나타나지 않았으며, 괴사율 또한 매우 낮은 수준인 것으로 조사되었다(Table 7). 따라서 ‘샤인머스캣’ 포도에서 발생하는 빈가지는 액아의 괴사보다는 낮은 화원기형성률에 기인하는 것으로 판단되었다.
Table 7.
Comparison of bud necrosis rate of ‘Shine Muscat’ grapevine according to growth regulators and light treatments in the growing period
| Treatment | Bud necrosis in Junez (%) | Bud necrosis in July (%) | Bud necrosis in August (%) | Bud necrosis in October (%) |
| Control | 1.67 | 2.54 ay | 3.39 a | 1.69 a |
| BAP | 1.67 | 4.24 a | 2.56 a | 3.36 a |
| SPD | 1.67 | 0.84 a | 5.88 a | 3.42 a |
| Long-Day | 1.67 | 1.68 a | 4.17 a | 0.86 a |
Cytokinin 처리시 꽃눈이 증가되었다는 이전의 보고(Koutinas et al., 2010)와 마찬가지로 BAP 처리시 화원기 형성이 가장 촉진된 것을 알 수 있었다(Table 8). 담배에 SPD 처리시 꽃눈이 증가하였다고 보고되었지만(Kaur-Sawhney et al., 1988), ‘샤인머스캣’ 포도의 SPD 처리는 생육기의 화원기 형성을 오히려 지연시켰으며, 휴면기에도 통계적 유의성은 없었지만 SPD 처리의 화원기형성률이 36.75%로 가장 낮은 것을 확인할 수 있었다. 따라서 BAP 처리가 액아의 크기 신장을 지연시켰으나 화원기 형성을 촉진한 반면 SPD 처리는 액아의 크기 신장은 촉진하고 화원기 형성은 지연시키는 결과를 보여, 같은 세포분열을 촉진하는 호르몬이라도 작용 부위가 생식기관과 영양기관으로 다를 수 있음을 보여주었다(Reece et al., 2011). 장일 처리의 경우, 생육기에는 화원기 형성을 늦추는 듯하였지만, 휴면기에는 무처리구와 비슷한 수준으로 화원기가 형성되었다(Table 8).
Table 8.
Comparison of inflorescence primordium formation rate of ‘Shine Muscat’ grapevine according to growth regulators and light treatments in the growing period
| Treatment |
Inflorescence primordium in Junez (%) |
Inflorescence primordium in July (%) |
Inflorescence primordium in August (%) |
Inflorescence primordium in October (%) |
| Control | 3.33 | 8.47 aby | 22.03 ab | 43.22 a |
| BAP | 3.33 | 14.01 a | 23.08 a | 47.06 a |
| SPD | 3.33 | 7.56 ab | 11.76 b | 36.75 a |
| Long-Day | 3.33 | 4.20 b | 17.50 ab | 41.38 a |
포도의 전정은 1, 2번 액아만 사용하는 단초전정, 1번 액아부터 6번 액아까지 사용하는 중초전정, 7번 액아 이상을 사용하는 장초전정으로 나뉘며, 액아는 형성되는 마디의 위치에 따라 충실한 눈의 형성 정도가 다르기 때문에(Khanduja and Balasubrahmanyam, 1972) 무처리구를 이용하여 각 전정법에 사용되는 액아의 화원기형성률을 비교하였다. 1, 2번 액아의 화원기 형성이 다른 위치에 있는 액아의 화원기 형성에 비해 현저히 낮은 것을 알 수 있었다(Fig. 2). 즉, ‘샤인머스캣’ 포도의 경우 중초 또는 장초전정을 하여야 빈가지 발생을 줄일수 있다고 생각되었다. 그러나 현재 대부분의 농가에서 단초전정으로 ‘샤인머스캣’을 재배하고 있어, 실제로 이용되는 1, 2번 액아에 대한 생장조절제와 빛 처리가 화원기 형성에 어떠한 영향을 미치는지 확인하였다. 1, 2번 액아의 화원기형성률을 처리구 별로 비교한 결과, BAP 처리에서 화원기 형성이 촉진되었다는 것을 확인하였다. SPD 처리의 경우 화원기 형성이 억제되었고, 장일 처리는 무처리와 비슷한 화원기 형성을 보였다(Fig. 3). 1, 2번 액아의 화원기형성률을 월별로 비교한 결과, BAP 처리가 화원기 형성을 가장 빠르게 촉진하였으며 장일 처리는 생육 후반기부터 형성을 촉진하는 것을 확인하여 각 처리가 화원기 형성에 미치는 영향은 1-10번 마디까지의 화원기 형성에 미치는 결과와 유사하였다. SPD 처리는 화원기 형성 속도가 가장 느렸고 형성률도 현저히 떨어졌다. 내재휴면이 끝난 후 3월의 화원기형성률은 모든 처리구에서 휴면기보다 낮아지는 것을 확인할 수 있었다. 이는 ‘캠벨얼리’ 포도를 대상으로 한 실험에서 내재휴면기와 내재휴면이 끝난 후 2월까지 겨울 동안에도 주아괴사, 부아발달, 화원기 형성 등이 계속 진행된다는 Lee(2015)의 보고와 마찬가지로 ‘샤인머스켓’ 포도에서도 겨울 동안 액아 내의, 주아, 부아, 화원기 형성이 계속 변화하기 때문인 것으로 생각된다. 내재휴면이 끝난 후에도 SPD 처리구는 다른 처리구에 비해 유의하게 낮은 화원기형성률을 보였다. 이를 통해 SPD 처리는 관찰 시기에 상관없이 화원기형성률을 억제하는 것으로 판단된다. BAP 처리, 장일처리 그리고 무처리의 화원기형성률은 휴면타파 후 비슷한 수준을 보이는 것을 확인하였다(Fig. 4). 내재휴면타파 후 1번, 2번의 액아의 종경은 SPD 처리 시 4.41mm로 가장 컸으며 BAP 처리 시 3.94mm로 가장 작은 것으로 확인되었다. 횡경 또한 유의성은 없었지만 SPD 처리 시 가장 크고 BAP 처리 시 가장 작은 것을 확인하였다. 이는 앞선 생육기와 환경휴면기의 1번 액아부터 10번 액아의 크기 생장 경향과 유사하였다. 눈 괴사율의 경우, SPD 처리 시 가장 낮은 수치를 보였지만 통계적 유의성은 나타나지 않았다(Table 9).
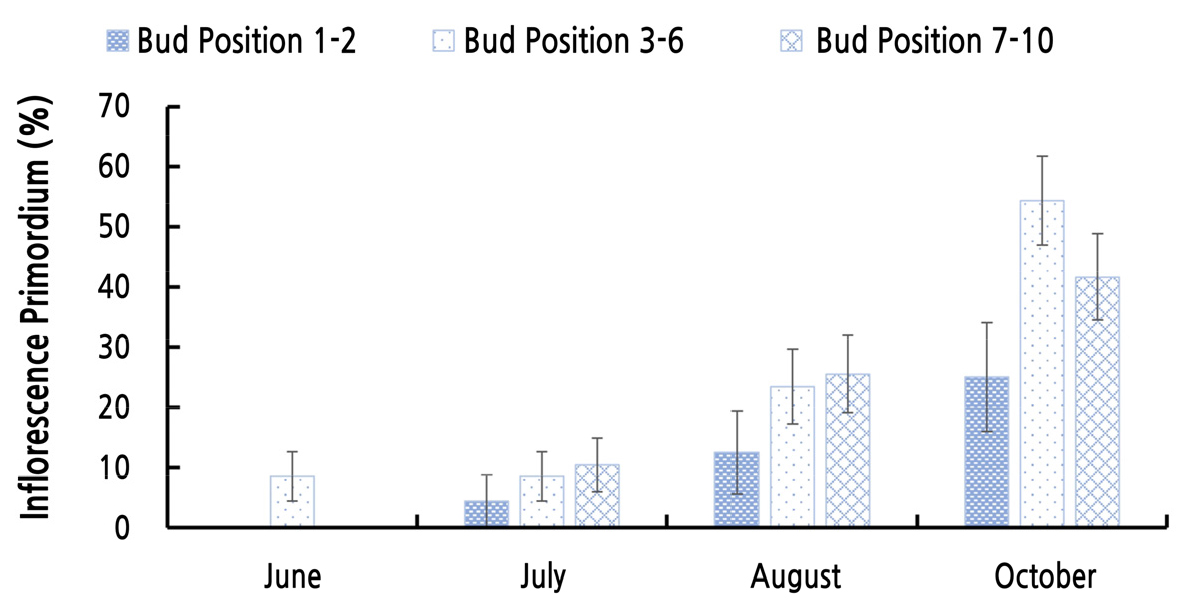
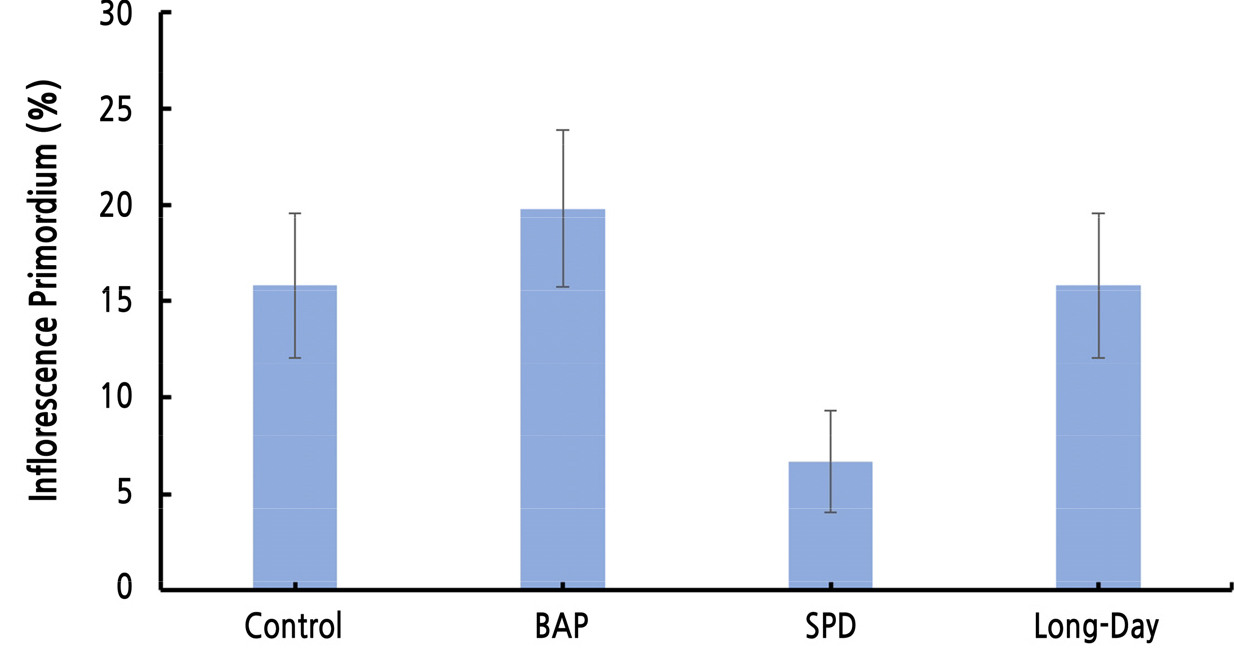
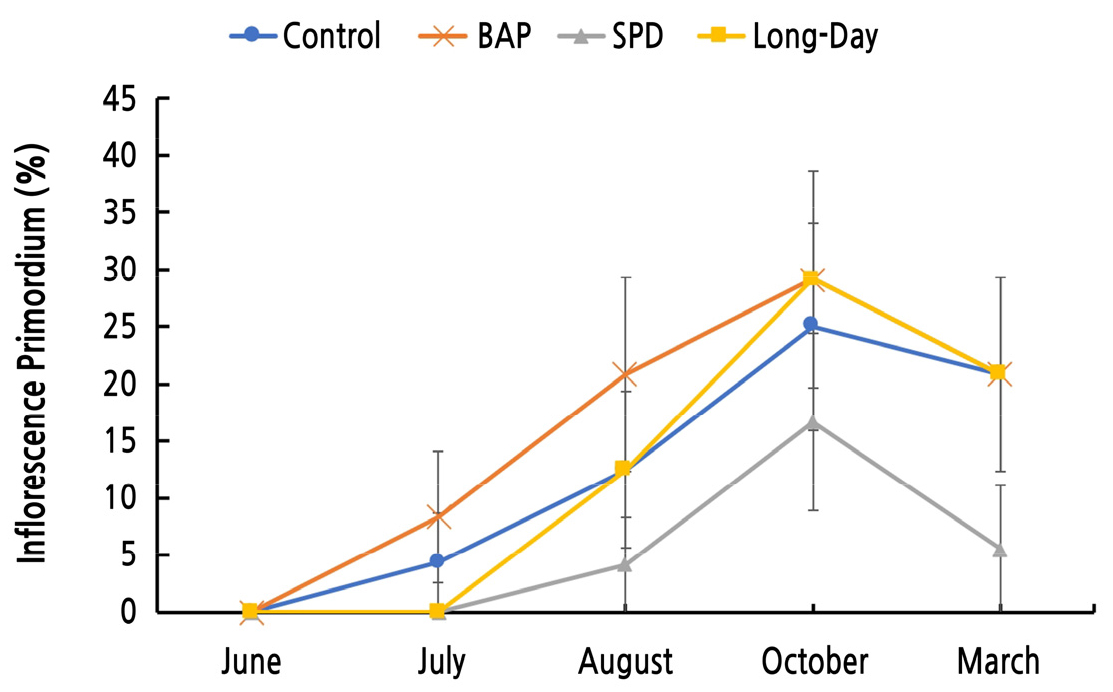
Table 9.
Bud length, bud diameter, and bud necrosis of 1st and 2nd buds of ‘Shine Muscat’ grapevine in March 2020 after growth regulator and light treatments in the growing period
| Treatment | Bud size | Bud necrosis (%) | |
| Bud length (mm) | Bud diameter (mm) | ||
| Control | 4.07 abz | 3.24 a | 8.33 a |
| BAP | 3.94 b | 3.36 a | 8.33 a |
| SPD | 4.41 a | 3.70 a | 5.56 a |
| Long-Day | 4.18 ab | 3.29 a | 8.33 a |
액아 발아 후 실제 포장에서의 꽃송이형성률(전체 신초 수 대비 꽃송이 수)은 휴면 타파 후 1번, 2번 액아의 화원기형성률과 마찬가지로 BAP 처리와 장일 처리는 무처리와 비슷한 수준의 꽃송이 형성률을 보였다. SPD 처리의 경우, 화원기형성률은 다른 처리에 비해 낮은 수치를 보였지만 꽃송이 출현율에서는 통계적 유의성은 나타나지 않았다(Fig. 5). 이는 내재휴면이 끝난 후 발아 전까지 계속적으로 화원기 분화가 이루어 졌기 때문으로 생각된다(Vasconcelos et al., 2009). 이를 통해 생육기의 BAP 처리와 장일 처리는 화원기 형성 속도에는 영향을 미치지만 형성률에는 큰 영향을 미치지 못하는 것으로 생각되며, SPD 처리는 화원기형성을 억제하는 것으로 판단된다.
두 시기별 실험의 결과, cytokinin이 꽃눈 형성과 개화에 관련되어 있다는 보고(Koutinas et al., 2010; Nambiar et al., 2012)와 마찬가지로 BAP 처리시 화원기 형성이 촉진되었다. 특히 내재휴면이 끝난 후 발아 전의 처리가 꽃송이 형성에 가장 효과적이었다. 생육기의 SPD 처리는 가지의 길이 생장과 액아의 크기 생장은 촉진하였지만, SPD 처리가 개화율과 꽃눈 형성을 증가시킨다는 이전의 보고(Kaur-Sawhney et al, 1988; Khezri et al., 2010; Qin et al., 2019)와는 다르게 환경휴면기의 SPD 처리는 화원기 형성에 영향을 미치지 않았으며, 생육기의 SPD 처리는 오히려 화원기 형성을 억제하였다. Koutinas et al.(2010)은 식물 지상부의 생장이 증대될수록 꽃눈 형성을 간접적으로 감소시킨다고 하였다. 따라서 SPD 처리는 ‘샤인머스캣’ 포도의 가지와 액아의 생장을 증대시킨 반면, 그 영향으로 액아의 화원기 형성이 억제되었다고 생각되었다. Buttrose(1969)는 포도의 장일처리가 화원기를 증가시킨다고 하였으나 ‘샤인머스캣’포도의 화원기 형성률에는 영향을 미치지 않았다. 따라서 ‘샤인머스캣’ 포도 액아의 화원기 형성을 촉진시켜서 빈가지 발생을 줄이기 위해서는 BAP 처리가 가장 효과적일 것으로 판단되었으며, 특히 환경휴면기에 200ppm의 BAP 농도에서 가장 효과적일 것으로 생각된다.
