

서 언
잔고사리과(Dennstaedtiaceae)의 돌토끼고사리[Microlepia strigosa (Thunb.) C. Presl]는 대한민국 원산 이며, 남제주의 바닷가 바위틈에서 자생하는 상록성 다년생 양치식물이다. 근경은 옆으로 뻗으며 지면 위로 올라오는 특성이 있다. 엽병은 30 ? 60cm이며 하부에 털이 많고, 엽신은 20 - 80cm의 예첨두형으로 2회 우상한다. 포자낭군은 거치 바로 아래의 가까운 축에 맺힌다. 연녹색의 어린잎은 관상가치가 우수하 여 남부지방에서는 노지에 조성된 암석원 등의 소재로 활용하고, 중부지방에서는 실내조경용 또는 분경 소재로 이용할 수 있다(KBIS, 2016).
한편 돌토끼고사리는 학술적, 생태적으로 중요도가 높은 식물구계학적 특정식물종 IV등급에 지정되어 있으므로(ME, 2007), 증식 및 이용 방안의 개발 등에 대한 연구의 필요성이 매우 높고 추후 수요가 증가할 것으로 예상되는 자생 양치식물 자원이다. 그러나 현재까지 돌토끼고사리에 대한 연구는 전무한 실정이며, 자생지 훼손으로 인한 대량 증식법 개발을 위한 체 계적인 증식 연구가 우선적으로 요구되고 있다.
종자식물군의 번식과는 달리, 대부분의 양치식물은 포자가 발아하여 전엽체를 형성하고, 전엽체에서 수정이 이루어져 포 자체가 형성되는 다소 복잡한 생활환을 가지고 있다. 따라서 양치식물의 증식에 있어서 생육단계별로 조직배양 기술을 적용 하는 것은 기본적인 발달생리의 기작을 해석하는데 매우 유용하다(Shin and Lee, 2009; Shin et al., 2010). 또한 특정 양치식물의 포자발아를 통해 생산된 전엽체에서 포자체로의 전환 및 포자체 육묘를 통한 규격묘의 생산은 경제적 가치를 지닌 식물 종의 산업화를 위한 대량증식의 실질적인 방안으로서 매우 중요한 과정이라 할 수 있다(Fernandez and Revilla, 2003). 따라서 양치식 물의 연중 대량생산을 위해서는 기내에서 포자, 전엽체 및 포자체 등 필요로 하는 특정 생육단계의 개체를 증식시킬 수 있는 방법이 요구될 수도 있다. 또한 생육단계별 조직배양 조건이 구명된다면 Park et al.(2015)와 Jang et al.(2016)의 연구에서와 같이 생물반응기 시스템에 접목함으로써 산업화가 촉진될 수 있을 것으로 생각된다. 따라서 본 연구에서는 돌토끼고사리의 포자 발아, 전엽체 증식, 포자체 형성 및 규격묘 생산을 위한 단계별 연구를 수행함으로써 효과적인 대량 증식법을 개발하고자 하 였다.
재료 및 방법
재료 및 방법
포자 채집
돌토끼고사리의 성체는 제주특별자치도 서귀포시 안덕면 일대에서 수집하여 충북대학교 농업생명환경대학 부속농장 부 지의 무가온 온실(70% 차광)에서 2년 이상 재배하였다. 이후 실험용 포자 채집을 위하여 2014년 07월 09일에 포자엽을 수확 한 다음 Shin and Lee(2009)의 방법으로 처리하였으며, 4 ± 1.0°C의 저장고에 건조상태로 보관하면서 연구에 사용하였다.
기내 포자의 발아
정선된 포자 50mg을 15mL 플라스틱 시험관에 넣고 1차 증류수에 24시간 동안 완전 침지한 다음 2,000rpm에서 3분간 원심 분리(MF 80, Hanil Sci., Seoul, Korea)하여 상징액을 제거하였다. 이후 1.4% sodium hypochlorite를 첨가하여 13분간 살균하였으 며, 동일한 방법으로 원심분리를 반복하여 멸균수로 5회 세척하였다. 또한 살균처리가 완료된 포자에 40mL의 멸균수를 첨 가함으로써 포자액을 제조하였다.
배지는 sucrose 3%, agar 0.8% 및 pH 5.8의 조건으로 통일한 MS배지를 1 / 8, 1 / 4, 1 / 2 및 1배의 농도로 조절한 것과 구성물 질의 함량 및 농도가 MS배지에 비해 현저히 낮은 Knop배지(Toole and Tottingham, 1918)를 조제하여 사용하였다. 이후 8.9mm 의 페트리접시에 각 30mL씩 분주하여 굳힌 다음 표면에 멸균수 1mL를 도포하였으며, 준비된 포자액을 1mL씩 접종하였다.
모든 발아실험은 4반복으로 수행되었으며, 접종이 완료된 페트리접시는 온도 25 ± 1.0°C, 광도 30 ± 1.0 μmol·m-2·s-1(16 / 8h) 인 배양실에 배치하였다. 이후 4일 간격으로 실체현미경(Eclipse E600, Nikon, Tokyo, Japan)을 이용하여 열개 및 발아한 포자 의 수를 단위면적에 대한 발아율로 계산하였으며, stereomicroscope(SZ51, Olympus, Tokyo, Japan)을 이용하여 발아된 포자 및 전엽체를 관찰하여 기관분화 등을 비교하였다.
기내 전엽체의 증식
2014년도에 채집한 포자를 기내 발아시켜 전엽체를 획득한 다음 2개월 간격으로 MS배지에 계대배양하여 실험재료로 사용하였다. 이때 MS배지의 sucrose와 agar의 농도는 각 3, 0.8%이며 pH는 5.8로 조절하여 조제하였다. 기내 전엽체 증식 연구는 Shin and Lee(2009)의 연구 방법을 응용하였다. 전엽체 증식에 적합한 배지를 선발하기 위하여 MS배지(sucrose 3%, agar 0.8%, pH 5.8)의 농도를 1 / 8, 1 / 4, 1 / 2, 1 및 2배로 조절하거나 Knop배지를 사용하였다. 이후 선발된 증식용 배지를 기본으로 하 여 배지 내 탄소원인 sucrose의 농도를 0, 1, 2, 3 및 4%로 달리하였다. 또한 활성탄(083006, Samchun pure chemical, Korea)이 전 엽체 배양에 미치는 영향을 구명하기 위하여 0, 0.2, 0.4 및 0.8%로 첨가한 배지를 조성하였다.
모든 처리는 4반복으로 수행되었으며, 조성된 각 배지에는 300mg의 전엽체를 메스로 다져서 접종한 다음 멸균수를 첨가 하였다. 접종이 완료된 배지는 온도 25 ± 1.0°C, 광도 43 ± 2.0μmol·m-2·s-1(16 / 8h)인 배양실에 배치하였다. 이후 8주 동안 배양 한 다음 전엽체의 생체중을 조사하였으며, stereomicroscope을 이용하여 전엽체의 발달과 형태형성을 관찰하였다.
기외 포자체 형성
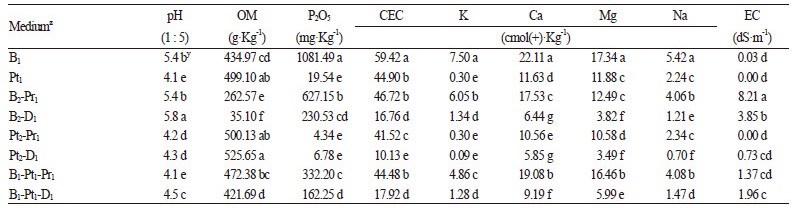
상기의 연구와 동일한 전엽체를 실험재료로 사용하였으며, Shin et al.(2010)의 연구에서와 같이 전엽체를 소독하였다. 기내 에서 증식된 전엽체로부터 대량의 포자체 형성에 유리한 배양토를 구명하고자 총 4가지 시판하는 인공토양을 단용하거나 비율별로 혼용한 8종류의 배양토를 조성하였다. 사용된 각 토양의 물리적 특성으로는 원예용상토(한아름상토 2호, (주)신성 미네랄, Korea)의 보수력이 70 ± 10%, 피트모스(SunShine, Sun Gro Horticulture, Canada)는 전공극률이 94.2 - 95.2%이었다. 펄라 이트(뉴펄샤인 2호, (주)지에프씨, Korea)는 유효수분율과 투수계수가 각 40%, 10-3cm·s-1 이상이고, 마사토(마사토 미립자, 김 해삼계마사, Korea)는 공시 입자크기가 2mm인 것을 사용하였다. 실험에 사용된 8가지 혼합토양은 Warncke(1986)의 방법으 로 상토의 화학성을 분석하여(serial no. 596 - 603) Table 1에 나타내었다.
이후 4각 화분(7.5 × 7.5 × 8.3cm, Cosmo corporation, USA)에 각 토양을 담아 평탄화하였으며, 전엽체 1g 당 25mL의 증류수 와 함께 hand blenders(V - 8000, Boowon, Korea)로 10초간 분쇄하여 준비된 화분에 균일하게 분주하였다. 처리가 완료된 화분 은 25L의 불투명 플라스틱상자[503 × 335 × 195mm, SPC532(DP - D), SH PLASTIC, Korea]에 담아 온도 25 ± 1.0°C, 광도 30 ± 2.0μmol·m-2·s-1(16 / 8h)인 배양실에 완전임의 배치하였다. 이후 재배기간 동안 플라스틱 박스 내 1cm 높이로 저면관수 하였 으며, 상부에 투명 유리판을 덮어 자체 습도(RH 72 ± 2.0%)를 유지하였다. 또한 전엽체가 발생하기 시작한 화분에는 1일 1회 두상관수를 실시하였다. 총 12주 동안 재배하였으며, 최종적으로 형성된 포자체의 수, 초장, 초폭, 엽장, 엽폭, 엽수, SPAD, 생 체중 및 건체중 등을 측정하였다.
포자체의 육묘
기내 배양을 통해 생산된 포자체를 삽목상자에 원예용상토를 담아 이식하였으며, 36L의 플라스틱 박스(System livingbox, Daewoo chemical, Korea) 바닥에 물을 일정량 채운 다음 삽목상자를 넣고 뚜껑을 닫아주었다. 이후 3주동안 실내에서 1차 순 화 후 90% 차광의 무가온 비닐온실에서 2차 순화하였으며, 이때 1일 2회 두상관수를 실시하였다. 이후 순화가 끝난 균일한 크기의 유묘를 실험에 사용하였다.
육묘용 플러그 트레이는 크기를 50, 72 및 105공 등 3종류로 달리하였으며, 토양은 상토와 마사토를 1 : 0, 2 : 1, 1 : 1, 1 : 2 및 0 : 1(v : v)의 비율로 혼합하여 5가지를 사용하였다. 각 처리는 20주씩 4반복으로 식재한 다음 70% 차광의 무가온 비닐온 실에 완전임의 배치하였으며, 2015년 7월 17일부터 2015년 11월 6일까지 16주 동안 재배하였다. 재배기간 동안 분수호스로 1 일 2회 두상관수 하였으며, 최종적으로 초장, 초폭, 엽장, 엽폭, 엽수, SPAD, 생체중 및 건체중 등의 생육반응을 측정하여 비 교분석하였다.
통계처리
모든 실험에서 조사된 측정값들에 대해 SAS version 9.3(SAS institute Inc., Cary, NC, USA)를 이용하여 평균과 표준오차를 구 하였으며, Duncan's multiple range test를 통해 유의성을 검정하였다( p < 0.05).
결과 및 고찰
결과 및 고찰
포자의 발아
배지의 종류 및 농도에 따른 돌토끼고사리 포자의 발아경향을 분석한 결과, 모든 배지에서 발아가 진행되었으나 발아개 시, 발아세 및 발아종료일 등 발아특성에는 차이를 보였다. 특히 Knop배지에서 발아개시 후 초기 발아세(0 - 8일)가 가장 높 았으며, MS계통의 배지에 비해 최종발아율에 먼저 도달하였다(Fig. 1A). 또한 배지의 영양분 및 sucrose의 농도가 낮은 배지 일수록 배지별 초기 발아세가 향상되었으나, MS계통에서는 MS배지에서 1 / 8, 1 / 4 및 1 / 2 MS배지에 비해 초기 발아세와 발아개시일이 저조하였다.
Fig. 1.
Effect of culture medium on spore germination in Microlepia strigosa (Thunb.) C. Presl. Vertical bars indicate standard error of the means (n = 4). Duncan’s multiple range tests at p < 0.05.
포자의 최종 발아율도 Knop배지와 1 / 8, 1 / 4 MS배지 등 영양분의 농도가 비교적 낮은 배지에서 높았으며, 모두 100%를 기록하였다(Fig. 1B). 또한 MS계통의 배지에서는 영양분 농도가 높아질수록 최종 발아율이 감소되었으며, MS배지에서 80.6%로 발아율이 가장 낮았다.
영양분의 농도가 상대적으로 낮은 배지에서는 삼투압에 의한 포자의 수분흡수가 촉진되기 때문에(Cox et al., 2003) 발아개 시에 유리한 조건이 조성될 가능성이 높다. 또한 포자도 종자와 유사하게 발아와 성장에 요구되는 일정량의 영양소를 함유 하고 있으므로(Whittier, 1975), 배지 내에 요구량 이상의 영양소 과잉은 본 연구에서와 같이 오히려 발아를 억제할 수도있 을 것으로 생각되었다.
포자발아 후 형성된 전엽체의 기관분화 정도를 관찰한 결과, 발아개시가 빠르고 발아율이 우수하였던 Knop배지에서는 날 개(wing)부분이 넓은 심장형의 전엽체가 발달하였다(Fig. 2A). 또한 Knop배지 내 전엽체의 상부에는 전엽체를 신장시키는 V 자형 정점(apical notch)이 밀집된 세포의 형태로 존재하였으며, 전엽체 뒷면에는 생식기관인 장정기(antheridium)가 날개부분 에 형성되었고 장란기(archegonium)가 가근(rhizoid)의 돌출부에서 관찰되었다.
Fig. 2.
Cultural responses and sexual organ formation in Microlepia strigosa (Thunb.) C. Presl prothalli cultured on Knop (A) and MS (B) media. an, antheridium; ap, apical notch; ar, archegonium; gp, germ filament; rh, rhizoid; sp, spatulate plate.
양치식물은 동일한 크기의 포자를 형성하는 동형포자형(homosporous)과 크기가 다른 소포자와 대포자를 형성하는 이형포 자형(heterosporous)으로 구분할 수 있다. 이형포자형은 소포자에서 웅성배우체와 정자가 발달되고 대포자에서는 자성 배우 체와 난자가 발달하여 성별이 다른 전엽체가 형성되는 반면, 동형포자형은 난자와 정자를 모두 생성하는 양성의 전엽체로 발달된다(Bold et al., 1987). 본 연구에서는 돌토끼고사리의 정상적으로 발달된 전엽체에서 장정기와 장란기가 동시에 관찰 되었으므로(Fig. 2A), 자생 양치식물인 도깨비고비, 참쇠고비(Jeong and Lee, 2006) 등과 같이 양성의 배우체를 발달시키는 동 형포자형 양치식물로 판단되었다.
한편 포자 발아율이 가장 낮았던 MS배지에서는 전엽체의 가장 초기단계인 필라멘트형(germ filament)과 다음단계인 말단 세포의 종적분열을 시작으로 형성되는 주걱형(spatulate plate) 전엽체가 관찰되었으므로(Fig. 2B), MS배지에서는 동일기간 동 안 Knop배지에 비해 정상적인 전엽체의 발달이 억제된 것으로 판단되었다 .
Chang et al.(2007)은 고란초과의 Drynaria fortunei 포자가 고농도의 MS배지에서 포자발아와 전엽체의 발달이 억제된 현상 을 배지 내 무기염류가 과다하여 발생한 것으로 분석하여, 본 연구와 유사한 결과를 보였다. 한편 Knop(1865)의 Knop배지는 예전부터 양치식물 포자발아에 통용되었는데, 본 연구에서도 MS배지에 비해 무기염류뿐만 아니라 탄소원의 함량도 절대적 으로 낮은 Knop배지에서 발아촉진 효과 및 심장형 전엽체의 발달을 유도하였으므로 돌토끼고사리 포자발아에 적절한 배지 조건으로 판단되었다.
전엽체의 증식
무기물 및 비타민의 농도를 달리한 배지에 전엽체를 분쇄하여 접종한 다음 8주간 배양한 결과, 포자발아를 위한 배지조건 과는 상반되는 결과를 나타내었다. 영양분의 함량이 높은 2X MS와 MS배지에서는 생체중이 각 59.8, 55.3배 증가하여 전엽 체 증식이 왕성하였으며, 유의적인 차이는 없었다(Fig. 3). 또한 MS계통에서는 배지 내 구성물질의 농도가 낮아질수록 전엽 체의 생체량이 유의적으로 감소되는 경향이었으며, Knop배지에서도 생육이 저조하였다.
Fig. 3.
Effect of culture medium on prothallus growth in Microlepia strigosa (Thunb.) C. Presl cultured for 8 weeks. Vertical bars indicate standard error of the means (n = 4). Duncan’s multiple range tests at p < 0.05.
원생소포자낭아강(leptosporangiate)에 속하는 양치식물의 전엽체는 일반적으로 영양물질의 농도가 낮은 배지에서 생육이 우수한 것으로 알려져 있으며(Fernsndez and Revilla, 2003), 자생 양치식물 중에서 고비의 전엽체는 Knop 또는 1 / 8 MS배지에 서 (Shin and Lee, 2009) 생육이 왕성한 것으로 보고된 바 있다. 반면 고사리(Shin et al., 2009)의 전엽체는 2X MS배지와 같이 영양물질의 함량이 비교적 높은 배지에서 생육 및 증식이 우수하여, 본 연구의 돌토끼고사리처럼 예외적인 결과를 보였다. 따라서 전엽체의 증식을 위한 영양분의 요구도에는 분류군에 따른 일정한 경향이 확인되지 않으므로(Shin et al., 2009), 전엽 체를 이용한 대량증식법의 개발을 위해서는 우선적으로 종에 따른 적정 배지를 선발하는 것이 반드시 선행되어야 할 것으 로 생각되었다.
배지별로 증식된 전엽체의 형태를 관찰한 결과, 무기물 및 비타민의 농도가 높은 2X MS와 MS배지에서 날개가 발달한 심 장형 전엽체로 발달되었으며, 모든 처리구에서 공통적으로 장란기 쿠션(archegonial cushion)이 관찰되었다(Fig. 4). 그러나 1 / 2, 1 / 4 및 1 / 8 MS배지에서는 심장형 전엽체의 전 단계인 주걱형 전엽체로 확인되어 발달이 저조한 것으로 판단되었다. 한 편 Knop배지에서는 전엽체의 색이 흐리고 육질이 얇으며, 일부 갈변된 부위가 관찰되었는데, 이는 무기물과 탄소원의 낮은 함량으로 인하여(Shin and Lee, 2009) 발생한 노화의 징후로 생각된다.
Fig. 4.
Cultural responses and sexual organ formation in Microlepia strigosa (Thunb.) C. Presl prothalli cultured on different media. A, 2X MS; B, MS; C, 1 / 2 MS; D, 1 / 4 MS; E, 1 / 8 MS medium; F, Knop medium. an, antheridium; ap, apical notch; cu, cushion; r, rhizoid.
한편 2X MS배지의 전엽체는 V자형 정점과 가근뿐만 아니라 쿠션부위에서 생식기관인 다수의 장정기가 형성되었다 (Fig. 4). 쿠션은 전엽체의 중심부에 발달되는 다세포 층으로써 짙은 녹색으로 나타나며, 생식기간이 발달할 수 있는 성숙한 전엽체에서 관찰되기 때문에(Shorina, 2001), 돌토끼고사리 전엽체의 생육 및 기관발달을 위해서는 2X MS 배지를 사용하는 것이 가장 효과적일 것으로 판단되었다.
2X MS배지를 기본배지로 하여 sucrose의 농도를 달리한 결과, 생체중 증가량의 차이가 확인되었으며, 특히 1, 2% 처리구에 비해 3, 4% 처리구에서 유의적으로 높은 생체중을 기록하였다(Fig. 5). 특히 기본 농도인 3% 처리구(control)에서는 무첨가구에 비해 6배 이상의 생체중이 증가한 것으로 조사되었다. 전엽체의 발달에서도 엽질 및 기관의 형성 정도 등의 차이가 발생하였는데, 특히 3% 처리구에서 가장 많은 장정기가 형성되었다(Fig. 6). 또한 0 - 3% 농도에서는 날개가 있는 심장형의 전엽체로 발달하였으나 4% 처리구에서는 주걱형의 불완전 전엽체가 형성되었다. 한편 sucrose의 농도가 높을수록 엽질의 색 이 짙어지는 경향이었는데, 이는 쿠션의 두께가 비대하였기 때문으로 생각되었다.
Fig. 5.
Effect of sucrose concentration on prothallus growth in Microlepia strigosa (Thunb.) C. Presl cultured for 8 weeks. Vertical bars indicate standard error of the means (n = 4). Duncan’s multiple range tests at p < 0.05.
Fig. 6.
Cultural responses and sexual organ formation in Microlepia strigosa (Thunb.) C. Presl prothalli cultured on 2X MS medium containing various concentrations of sucrose. A, no treatment; B, 1%; C, 2%; D, 3%; E, 4%. an, antheridium; ap, apical notch; cu, cushion; r, rhizoid.
2X MS배지에 0.2 - 0.8%의 활성탄을 첨가한 결과, 무첨가구에 비해 전엽체의 생육이 유의적으로 억제되었으며, 생체중 증가량은 모든 활성탄 첨가구에서 농도에 관계없이 3.1 - 3.4g 감소되었다(Fig. 7). 전엽체의 형태를 관찰한 결과, 활성탄의 농도 가 높을수록 쿠션부위가 비정상적으로 두꺼워져 짙은 색을 나타내었으며, 생식기관인 장정기의 존재가 확인되지 않았다 (Fig. 8).
Fig. 7.
Effect of activated charcoal concentration on prothallus growth in Microlepia strigosa (Thunb.) C. Pres
Fig. 8.
Cultural responses and sexual organ formation in Microlepia strigosa (Thunb.) C. Presl prothalli cultured on 2X MS medium containing various concentrations of charcoal. A, no treatment; B, 0.2%; C, 0.4%; D, 0.8%. an, antheridium; ap, apical notch; cu, cushion; r, rhizoid.
일반적으로 기내에서 양치식물의 배양 시 sucrose는 전엽체의 생육을 돕는 것으로 알려져 있으며(Fernandezd와 Revilla, 2003), sucrose의 결여는 광합성을 제한하여 전엽체의 재생율을 감소시킬 수 있다(Ambrosio and Melo, 2004). 본 연구에서도 배 지 내 3%의 sucrose를 첨가하였을 때 돌토끼고사리 전엽체의 생육 및 발달에 긍정적인 효과가 확인되었으며, '이는 Blechnum spicant(Fernandez et al., 1997)에서 보고된 결과와 유사하다.'
활성탄은 배양된 조직이나 배지 내 sucrose의 고압살균을 통해 발생하는 5 - HMF(hydroxymethyl furfural) 등의 독성물질을 흡착하여 배지의 오염을 방지하는 역할을 한다. 또한 배지 내에서 초과된 옥신이나 배양된 조직에서 발생한 에틸렌 을등 흡 수하는 등(Moshkov et al., 2008) 다양한 이점을 갖고 있으나, 본 연구에서는 전엽체의 생육 및 발달에 대한 활성탄의 긍정적인 효과를 확인할 수 없었다.
연구의 결과, 돌토끼고사리 전엽체를 기내에서 대량 증식하고자 할 때는 sucrose 3%의 2X MS배지에 배양하는 것이 정상 적인 발달과 생체중 증가에 효과적이며, 활성탄은 첨가하지 않는 것이 좋을 것으로 판단된다 .
포자체의 형성
기외에서 전엽체로부터 포자체를 유도하기 위해 조성된 8가지 배양토의 이화학적 성분을 분석한 결과, pH는 피트모스가 함유된 모든 배양토에서 4.1 - 4.5 수준의 산성을 띄었으며, 상토 : 마사토 2 : 1(v : v) 혼합토양에서 pH 5.8로 가장 높았다(Table 1). 유기물 함량(OM)은 상토:마사토 2 : 1(v : v) 혼합토양에서 가장 높았으나, 오산화인(P2O5) 함량은 상토 단용 처리구에서 상 대적으로 가장 많았다. 한편 K, Ca, Mg 등의 무기물 함량 및 양이온 치환용량(CEC)은 상토 단용에서 유의적으로 가장 많았 으며, 전기전도도(EC)는 상토:펄라이트 2 : 1(v : v) 혼합토양에서 8.21dS·m-1로 가장 높았고, 배양토 내 피트모스의 비율이 높 을수록 전기전도도가 낮은 것으로 나타났다. 이러한 결과는 토양 입자가 작을수록 전기적 인력이 강해여 토양 내 수의분 무 기물 등 식물이 이용할 영양성분의 함량이 많을 뿐만 아니라 전기전도도도 높아질 수 있으나, 본 연구에서 분석한 바와 같이 토양 내 수분이 아닌 입자 자체의 전기전도도는 낮을 수도 있음을 시사한다. 또한 양치식물의 전엽체로부터 포자체의 형성 을 유도할 때 사용되는 토양의 입자, 공극률, 용기용수량, 전기전도도 등의 물리성과 화학성이 원예작물의 생육반응과 같이 복합적으로 관여함(Choi et al., 2011)을 확인할 수 있었다.
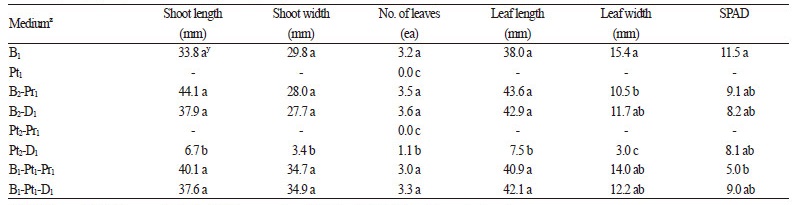
돌토끼고사리의 포자체를 대량으로 발생시키기 위하여 분쇄한 전엽체를 혼합비율을 달리한 배양토에 분주한 다음 12주 동안 재배하였다. 연구의 결과, 상토 : 피트모스 : 마사토를 1 : 1 : 1(v : v : v)로 혼용한 토양에서 포자체의 발생량이 1.75개 / cm2로 가장 많았으며, 상토 : 피트모스 : 펄라이트 1 : 1 : 1(v : v : v) 혼합토양, 상토:펄라이트 2 : 1(v : v) 혼합토양에서 각 1.07, 0.43개 / cm2로 조사되었다(Table 2). 한편 포자체 발생량이 가장 많았던 상토 : 피트모스 : 마사토 1 : 1 : 1(v : v : v) 혼합토양 에서 초장, 초폭, 엽수 및 엽장 등의 생육도 유의적으로 우수한 수준이므로(Table 3), 포자체 발생에 효과적인 배양토로 판단 되었다.
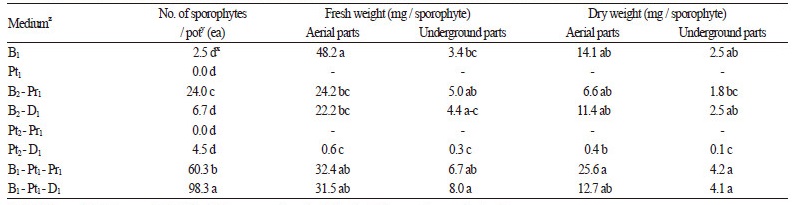
배양토 배합 시 입자의 크기를 달리하여 공극률, 통기성 및 보수성 등을 조절함으로써 작물생육에 유리한 토양환경을 조 성할 수 있다(Choi et al., 1997). 자생 양치식물의 경우 대체로 지생종은 원예용 상토 단용, 착생종은 질석 또는 펄라이트를 혼 합하여 통기력을 높인 토양에서 포자체의 형성이 왕성하였다(Lee, 2001). 본 연구의 돌토끼고사리도 상토에 피트모스와 마 사토를 동일한 비율로 혼합한 토양에서 포자체 발생량 및 생육이 우수하였는데, 이는 근경이 지면 위로 돌출하는 착생종(KBIS, 2016)의 생육환경과도 유사하였기 때문으로 생각된다.
한편 원예상토와 피트모스의 각 단용 및 피트모스의 첨가비율이 높은 토양에서는 전엽체가 재생되었음에도 불구하고 포 자체 발생이 억제되는 경향을 보였다(Table 2). 이는 재배기간 동안 유지된 저면관수 조건이 원예용 상토나 피트모스의 단용 처리구에 수분과다로 인한 통기성의 저조를 야기하여 포자체의 형성 및 뿌리발달에 악영향을 미쳤기 때문으로 생각된다. 또 한 피트모스의 단용 및 첨가비율이 높은 배양토에서 토양 입자의 전기전도도가 상대적으로 낮았던 것도 포자체 발생 억제 의 원인으로 생각된다(Table 1). 피트모스는 플러그 육묘용 혼합상토의 주재료로서 다량의 유기물을 함유하고 있으며, 양이 온 치환능력과 보비력이 우수하지만(Argo, 1998), 평균 pH가 3.8 ± 0.94인 강산성이므로(Shin et al., 2012) 사용시 주의를 요한 다. 한편 국내에 유통되는 피트모스는 대부분 수입산으로 원산지에 따라 부숙도에 차이가 발생하여 작물생육에 악영향을 줄 수 있으므로(Nelson, 2003), 돌토끼고사리의 포자체 형성에 있어서 다량의 사용은 피하는 것이 좋을 것으로 판단된다 .
포자체의 육묘
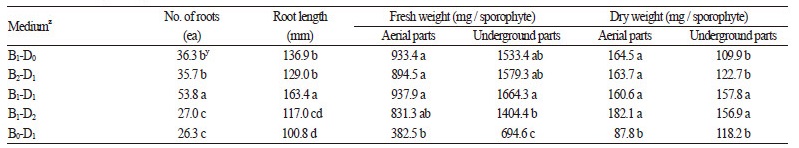
상토와 마사토의 비율을 달리한 육묘용토에서 순화를 마친 돌토끼고사리 유묘를 식재하여 16주 동안 재배한 결과, 상토 가 함유된 토양조건에서 공통적으로 지상부의 생육이 우수한 경향이었으며, 특히 상토 : 마사토 2 : 1(v : v) 혼합처리구에서모든 지상부의 생육상태가 타 처리구에 비해 가장 왕성하였다(Table 4). 지하부 생육을 분석한 결과, 상토의 비율이 50.0%인 토양조건에서 뿌리 발생량이 53.8개로 가장 많았으며, 뿌리의 신장도 활발하였다(Table 5). 또한 상토의 비율이 낮을수록 지 하부 생육은 유의적으로 억제되었으며, 마사토 단용 처리구에서는 근수와 근장이 각 26.3개, 100.8mm로 상대적인 저조를 보 였다. 따라서 육묘용토는 동일 재배기간 동안 지상부 생육이 가장 양호하고 지하부 생육에 장해가 없다고 판단되는 상토 : 마사토 2 : 1(v : v)이 적합할 것으로 생각된다.
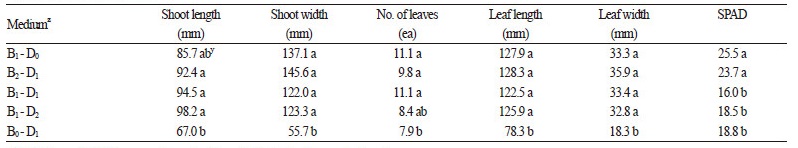
한편 상토의 비율이 50% 이하로 낮아질 경우 SPAD값이 유의적으로 감소되었는데(Table 4), 이는 토양 내 질소의 함량이 잎 의 건물중, 면적 및 엽록소의 함량에 적접적으로 관여하기 때문이며(Wang et al., 2012), 육묘용토 내 상토의 비율은 질소성분 의 함량을 좌우함으로써 포자체의 생육 차이를 야기할 수 있는 요인으로 작용하였기 때문으로 생각되었다.
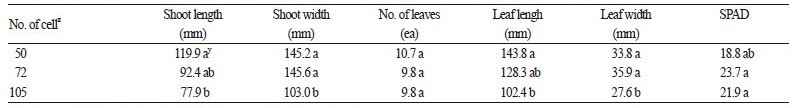
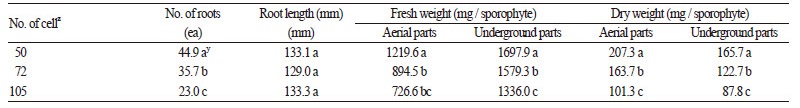
선발된 육묘용토[상토 : 마사토 2 : 1(v : v)]를 사용하여 트레이 크기별 돌토끼고사리 유묘의 생육반응을 비교 · 분석한 결 과, 50공에서 초장과 엽장이 유의적으로 가장 길었으며 공의 크기가 클수록 지상부의 생육에 유리한 조건이었다(Table 6). 지 하부 생육에서도 50공에서 뿌리 발생량이 44.9개로 가장 많았으며, 72, 105공 순으로 유의적인 감소세를 보였다(Table 7). 그 러나 뿌리의 길이는 모든 처리구에서 129.0 - 133.3cm로 유의적인 차이가 없었는데, 이는 재배기간 동안 뿌리신장이 포화생 육을 하였기 때문으로 생각된다. 한편 트레이 크기에 따른 생체량도 생육이 우수하였던 50공에서 가장 많았으며, 트레이의 크기가 클수록 생체량이 유의적으로 감소되었다. 돌토끼고사리는 지하경이 발달하여 지면을 피복하기 때문에(KSIB, 2016), 유묘 단계에서 지하부의 생육을 좋게 하는 것은 차후 피복용 소재로서 이용에 유리할 수 있다. 따라서 돌토끼고사리 육묘 시 지하부의 생육뿐만 아니라 지상부 생육을 최대화 하기 위해서는 50공 플러그 트레이를 사용하는 것이 효과적일 것으로 판단 된다.
본 연구의 결과, 돌토끼고사리의 번식을 위해서는 포자, 전엽체 및 포자체의 각 생육단계에서 요구되는 증식조건이 다양하였으며, 본 연구의 결과를 바탕으로 균일한 묘의 대량증식을 위한 체계적인 생산시스템의 구축이 개발될 수 있을 것으로 기대된다.
