

Introduction
Materials and Methods
Plant Material and Culture Conditions
Experimental Design
Histological Observation
Physiological Index Determination
Statistical Analysis
Results
Discussion
Introduction
Tree peony (Paeonia suffruticosa Andrews) is an ornamental plant that originated in China and is known for its beautiful flowers. This species has high ornamental and horticultural value (Bao, 2003). As the demand increases for cultured tree peony, the development of a sustainable tree peony propagation system is paramount. Tree peony is typically propagated by grafting the desired aerial cultivar onto rootstock from a certain species. However, Paeonia rootstocks are generally characterized by insufficient lateral root development and low root quality (Liu et al., 2013). Additionally, during transplanting, tree peony seedlings always suffer from mechanical root damage. Studies have found that cutting roots can promote lateral root development, increase the height of the plant, improve plant quality, facilitate lifting of seedlings and transplanting, shorten the recovery period, and increase the survival rate (Zou et al., 2006; Ma et al., 2009; Wang et al., 2010). However, the mechanism behind lateral root development in tree peony has not been studied. Therefore, knowing how to optimize the development of the root structure of tree peony rootstock is particularly important. Previous studies have found that the root development of 1-year-old seedlings or first-class fleshy roots can be improved by root pruning (Liu et al., 2013). After root cutting, the growth of the main root was inhibited, yet growth of lateral roots and root activity were improved significantly in rice (Oryza sativa) (Liu et al., 2014), Yunnan pine (Pinus yunnanensis) (Zhao et al., 2010), and grape (Vitis vinifera) seedlings (Zhang and Zhao, 2010). Additionally, the survival rate of in vitro transplants of grape plantlets was increased (Zhang and Zhao, 2010). Interestingly, the position of the cutting alters the effect on rooting (Zou et al., 2006).
Studies have shown that root pruning can alter the activities of oxidase-related enzymes, such as peroxidase (POD), superoxide dismutase (SOD), polyphenol oxidase (PPO), and indole acetate oxidase (IAAO) (Zhao et al., 2010; Yang et al., 2011), and alter the levels of endogenous hormones, such as indole acetic acid (IAA), zeatin (ZT), gibberellin (GA3), and abscisic acid (ABA) (Chen et al., 2014). These hormones work together to regulate root growth and development. Therefore, these related activates need to be measured during an investigation into the effects of root pruning on the early development of the root and to optimize the development of tree peony roots and improve rooting quality.
In this paper, the tree peony cultivar ‘Fengdanbai’ was used to study the changes in anatomical structure, morphology, and physiological and biochemical indexes during root development following different types of pruning of young roots from seedlings. The aim of this study was to develop an optimal pruning treatment during early development of the root architecture of seedlings by studying the development of the tree peony root system.
Materials and Methods
Plant Material and Culture Conditions
Mature seeds of the tree peony (Paeonia suffruticosa) variety ‘Fengdanbai’ were obtained from the Luoyang Shenzhou Tree Peony Garden. The seeds were sanded with sterilized river sand, and then placed in a ventilated shade house at 10–20°C. Water was added every 2–3 days (d), and the layer (Layer with gauze) was changed every 5 d. Pests and diseases were monitored during incubation in the sand.
Experimental Design
Pruning roots of the same length. roots were pruned by either 1/5, 1/3, 1/2, or 3/4 of their length or were left unpruned as a control when more than 50% of the seedling roots reached 2 cm in length. The materials were examined at 0, 1, 2, 3, 4, 5, 6, and 7 d after pruning. Three samples were randomly selected at the same time every day for measuring root length and counting the number of lateral roots. Lateral root emergence was determined by observation with the naked eye.
Pruning roots of different lengths. The above experiment showed that pruning roots by 1/3 was a promising treatment. Therefore, in this experiment, roots of different lengths were pruned by 1/3 of their length. Roots 1, 2, 3, or 4 cm (± 0.5 cm each) in length were pruned by 1/3 when more than 50% of the seed roots reached a 2 cm uniformity. The root length and number of lateral roots of 3 randomly selected samples were quantified at 0, 1, 2, 3, 4, 5, 6, and 7 d. Roots left unpruned were used as the control treatment.
The following calculations were performed:
Growth of the primary root (cm) = main root length at 7 d – main root length at 0 d.
Lateral rooting rate (%) = number of seedlings with lateral roots/total number of test seedlings × 100.
Average number of lateral roots (strip) = total number of lateral roots in the sampled seedlings/total number of seedlings sampled.
Histological Observation
Histological changes of the lateral roots were obtained by paraffin sections (Schleicher, 1951).
The tissues were fixed with 38% formalin or other fixatives for a week at room temperature. The fixed tissues were embedded in paraffin over a total of 34.5 hours: 50% ethanol, two changes, 1 hour each; 70% ethanol, two changes, 1 hour each; 85% ethanol, two changes, 1 hour each; 95% ethanol, two changes, 1 hour each; 100% ethanol, four changes, 0.5 hour each; xylene and 100% ethanol, one change, 2 hours; xylene (i.e., Clear Rite 3), three changes, 50 minutes each; paraffin wax (58–62°C), one change, 12 hours; paraffin wax (58–62°C), two changes, 4 hours each; and then the root sections were made into wax blocks Sections were cut at 20 mm (15 mm is commonly used) using the KD 202-A rotary slicer. The paraffin ribbon was placed into a water bath at 30–35°C. Sections were mounted onto slides, air-dried for 30 minutes, and then baked in an oven at 30–35°C overnight. The paraffin was removed in one change of xylene, 15–25 minutes. The tissues were hydrated in 2 changes of 100% ethanol for 2 minutes each, and then 95% ethanol for 1 minute. The samples were double-stained with Safranine and Fast Green and then rinsed in distilled water. The samples were observed and photographed using an Olympus optical microscope (Motic SZM171).
Physiological Index Determination
The histological observation determined that the key period of rooting was 3 to 5 d after root pruning. Therefore, relevant physiological indicators were determined 3 or 5 d after the optimal root pruning treatment. Root vigor was determined using the TTC (2'3'5-Triphenyte-trazoliumchloride) method (Li, 2000); peroxidase (POD) activity was determined using the guaiacol method (Li, 2000), superoxide dismutase (SOD) activity was determined by the NBT method (Zhao et al., 1998; Li, 2000); polyphenol oxidase (PPO) activity was assayed using the method described by Zhu et al. (1990); and indole acetate oxidase (IAAO) activity was assayed as described by Zhang (1990). The levels of the endogenous hormones indole acetic acid (IAA), zeatin (ZT), gibberellic acid (GA3), and abscisic acid (ABA) were determined by liquid chromatography-mass spectrometry. Fresh roots from the control group and the root pruning treatment group were frozen in liquid nitrogen, ground with glass beads, and further processed by liquid chromatography-mass spectrometry (Gou et al., 2010).
Statistical Analysis
The test data was processed and analyzed by Excel 2003 and DPS 7.05 software.
Data were subjected to one-way analysis of variance. Significant differences between means were tested using Duncan’s post-hoc test (p < 0.05). The test data was processed and analyzed by Excel 2003 (Microsoft) and Data Processing System (DPS) 7.05 software.
Results
The radicle of tree peony responded to early root pruning. The anatomical structure of the root primordium during formation and adventitious root emergence through the epidermis showed significant differences after pruning treatment (Fig. 1). Three days after pruning, lateral root primordium began to form (Fig. 1A and 1B). After 4 days, the growth points and root crowns were differentiated (Fig. 1C). After 5 days, the adventitious roots broke through the epidermis (Fig. 1D). Therefore, lateral roots emerged 3–5 days after root cutting. In the unpruned roots, lateral roots did not form at this time.
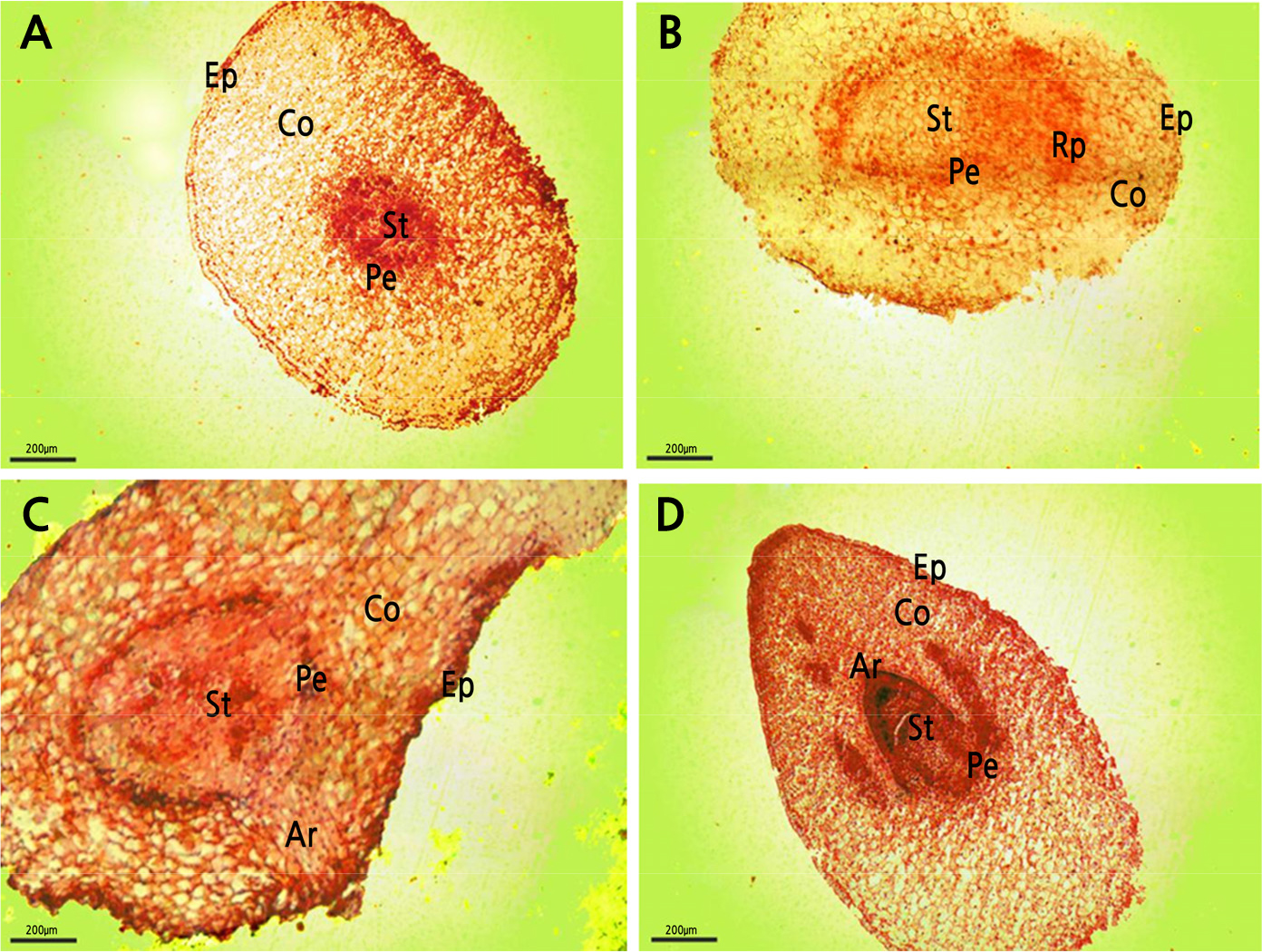
Fig. 1.
Root primordium at 0, 3, 4, and 5 d after root pruning. A: root pruning transverse section 0 d (100x); B: root pruning transverse section 3 d (100x); C: root pruning transverse section 4 d (100x); D: root pruning transverse section 5 d (100x). Co: cortex; Ep: epidermis; Rp: adventitious root primordium; Ar: adventitious root; Pe: pericycle; St: stele.
Seven days after pruning, the length of the primary roots increased. The greater the amount of pruning, the greater the growth after 7 days (Table 1). For instance, in roots that were pruned by 3/4 of their initial length (2 cm), the root grew 1.56 cm in 7 days, while the 2 cm roots pruned by only 1/5 grew less than 1 cm in 7 days. The unpruned roots showed the greatest growth (2.95 cm for those starting at 2 cm and 4.98 cm for those starting at 3 cm).
Table 1.
Root growth and lateral root emergence of tree peony seedlings 7 days after removing the end of the radicle
|
Average initial root lengthz (cm) |
Root pruning treatmenty |
Primary root growth (cm) |
Average number of lateral rootsx |
Lateral rooting rate (%) |
Earliest lateral root (d) |
| 2 | 1/5 | 0.94 ± 0.03 b | 0.96 ± 0.69 a | 60 | 4 |
| 1/3 | 1.02 ± 0.39 c | 1.12 ± 0.31 ab | 80 | 5 | |
| 1/2 | 1.07 ± 0.11 c | 0.42 ± 0.21 ab | 60 | 5 | |
| 3/4 | 1.56 ± 0.04 c | 0.54 ± 0.28 ab | 60 | 6 | |
| 0 | 2.95 ± 0.04 a | 0 | - | - | |
| 3 | 1/5 | 0.59 ± 0.03 bc | 1.44 ± 0.57 a | 80 | 4 |
| 1/3 | 0.50 ± 0.04 c | 2.34 ± 0.43 b | 90 | 4 | |
| 1/2 | 0.72 ± 0.03 bc | 0.84 ± 0.33 b | 70 | 4 | |
| 3/4 | 1.09 ± 0.02 b | 0.84 ± 0.48 b | 60 | 6 | |
| 0 | 4.98 ± 0.24 a | 0 | - | - |
The number of lateral roots negatively correlated with the length of root pruned. That is, the greater the amount of pruning, the fewer the number of lateral roots that were observed 7 days later. For both initial root lengths (2 and 3 cm), the greatest number of lateral roots (1.12 and 2.34, respectively) and highest rooting rate (80% and 90%, respectively), occurred following pruning the root by 1/3. No lateral roots were visible in the unpruned roots.
The earliest lateral root was observed 4 days after pruning by 1/5 (for roots initially 2 cm long) or by 1/5, 1/3, or 1/2 (for 3 cm long roots). While the roots pruned by 1/5 grew lateral roots faster, the root system was relatively slender. Pruning the roots by 1/2 or 3/4 produced lateral roots relatively late (at 5 or 6 days), and at a lower rate. Considering the length of the rooting period, the number of lateral roots emerged, and the degree of root thickening (Fig. 2), the 1/3 root pruning treatment was chosen as the best treatment.
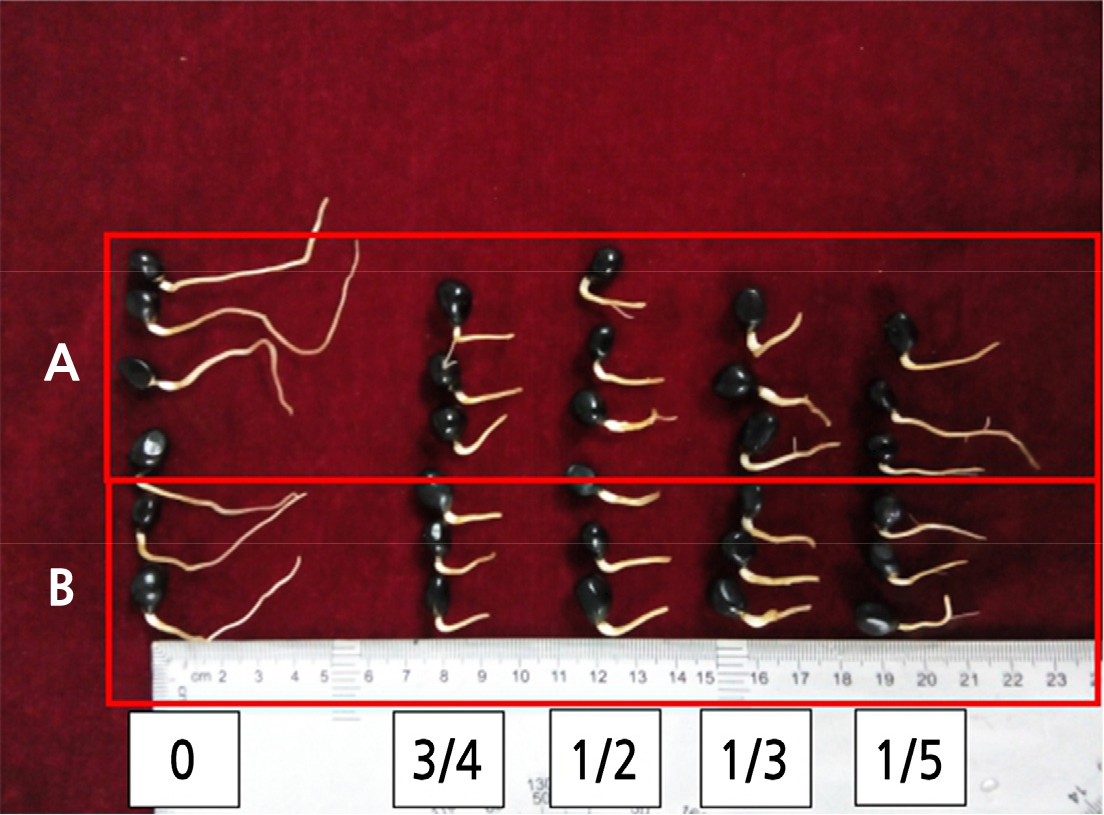
Fig. 2.
The different root pruning treatments of roots of length 2 and 3 cm after 7 d of the radicle growth. A: root length 3 cm; B: root length 2 cm. The 0, 3/4, 1/2, 1/3, or 1/5 in the bottom of the figure means that roots were pruned by 0, 3/4, 1/2, 1/3, or 1/5 of the radicle length, respectively (0 means unpruned).
The initial root length significantly affected the final root length and lateral root emergence when roots were pruned by 1/3 (Table 2). All pruned roots grew less than the unpruned roots, regardless of their initial length. Among the pruned roots, there was variation in how much they grew after pruning, with the most length added to roots that were initially 3 cm in length.
Table 2.
Root growth and lateral root emergence of tree peony seedlings 7 days after removing a third of the radicle
|
Average initial root lengthz (cm) |
Root pruning treatmenty |
Primary root growth (cm) |
Average number of lateral rootsx |
Lateral rooting rate (%) |
Earliest lateral root (d) |
| 1 | 1/3 | 0.70 ± 0.08 bcd | 0.18 ± 0.16 bc | 30 | 6 |
| 2 | 0.65 ± 0.04 bcd | 0.64 ± 0.29 b | 40 | 5 | |
| 3 | 0.85 ± 0.12 a | 1.07 ± 0.16 a | 60 | 4 | |
| 4 | 0.77 ± 0.06 b | 0.83 ± 0.55 a | 50 | 2 | |
| 1 | 0 | 0.82 ± 0.07 bcd | 0 | - | - |
| 2 | 1.00 ± 0.08 bc | 0 | - | - | |
| 3 | 1.36 ± 0.21 d | 0 | - | - | |
| 4 | 1.68 ± 0.10 cd | 0 | - | - |
None of the unpruned roots showed lateral root growth after 7 days. For the pruned roots, the average number of lateral roots increased with initial root length. The greatest number of average lateral roots (1.07) was observed in roots that were initially 3 cm before the 1/3 pruning. The lateral rooting rate also increased with increasing initial root length. The highest lateral rooting rate (60%) occurred for roots that were initially 3 cm in length, followed by those that were 4 cm (at 50% rooting rate). The time to the first lateral root decreased with increasing initial root length. Roots that were 4 cm before pruning formed lateral roots earlier (2 d), followed by those that were 3 cm (4 d).
The lateral roots that grew early (4 cm roots following 1/3 root pruning) were slenderer and more occurred at a lower rate than the roots that emerged from primary roots that were initially 1 and 2 cm in length (Fig. 2). The thicker lateral roots occurred slightly later (4 days after pruning) but at a higher rate. Taking the data comprehensively, the best combination was considered to be an initial root length of 3 cm with the 1/3 root pruning treatment. As a measure of the vitality of the tree peony roots, TTC was used to measure cellular respiration (Fig. 3). The TTC reduction activity of 3-cm roots pruned by 1/3 was higher than that of the unpruned roots over the 7-day period following pruning. The activity initially rose, peaking on the 3rd day, and then remained high during the critical rooting period over days 3 to 5. The differences between the pruning treatment and unpruned roots were significant (Fig. 3). The pruning treatment significantly improved root vitality during the lateral root formation period.
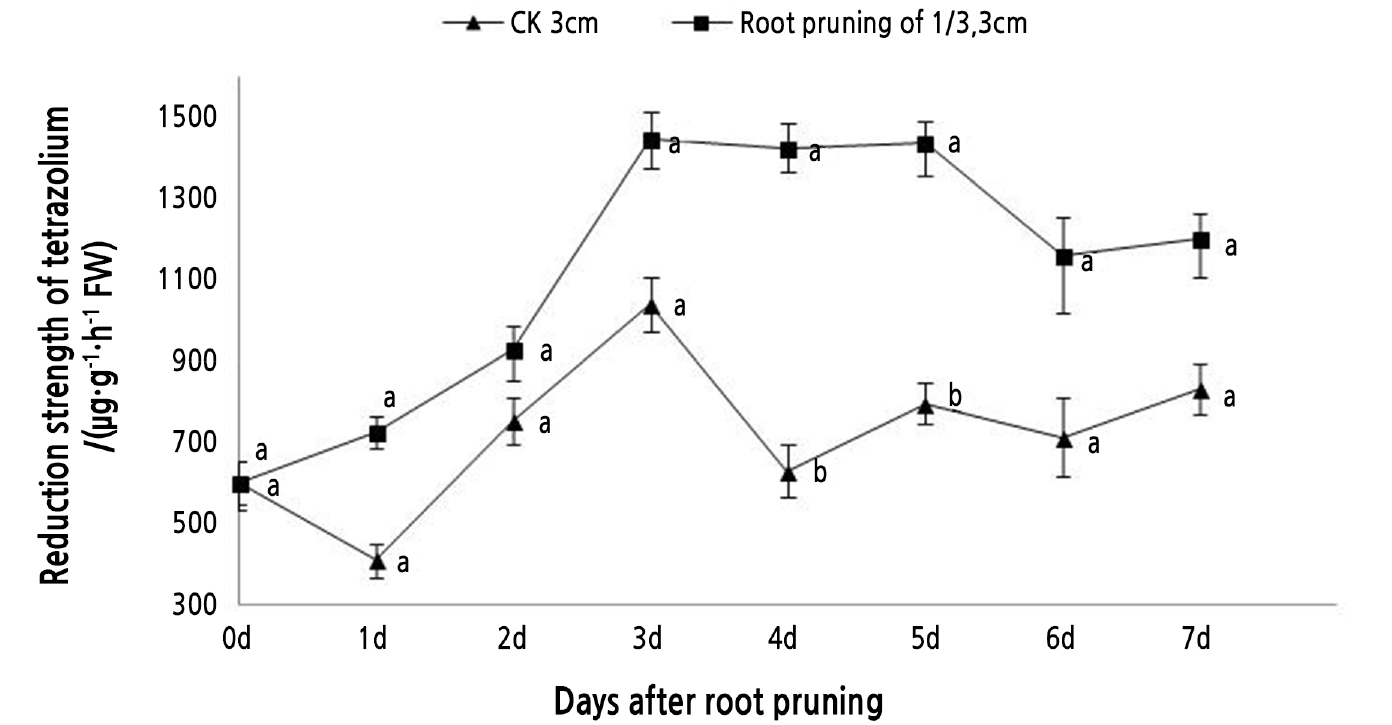
Fig. 3.
Tree peony seedlings with 3-cm long roots were pruned by 1/3. TTC reduction activity was measured over 7 days. Vertical bars indicate standard error. Each value is the mean obtained from replicates. CK 3 cm indicates the control tree peony seedlings with 3-cm long roots that were unpruned. Values on the same day with the same letter are not significantly different between pruned and unpruned roots by Duncan’s multiple range test at p < 0.05 (n = 3).
Antioxidant activity was measured over 5 days in 1/3-pruned and unpruned roots from seedlings with roots initially 3 cm in length (Fig. 4). The activity of POD generally increased over time in the pruned roots, but remained flat in the control (Fig. 4B). During root primordium formation (3–5 d), the POD activity in the pruned roots rapidly increased and was significantly higher than that of the control (Fig. 4B). On day 5, the POD activity in the pruned roots was 3.95 times that of the control. PPO activity did not show significant differences between pruned and unpruned roots (Fig. 4C). In the 1/3-pruned and unpruned roots, the levels of PPO activity fluctuated. IAAO activity showed an upward trend in both the pruning treatment and the control (Fig. 4A). However, there was an earlier peak 12.71% higher at 4 d in the pruned roots compared to the control (which peaked at 5 d). IAAO activity levels were not significantly different between the treatment and control 2 or 3 days after pruning. Root pruning significantly increased SOD activity 2 and 5 days after pruning (Fig. 4D). After the formation of the root primordium, the SOD activity in the pruned roots decreased, while the levels in the control first increased and then decreased (Fig. 4).
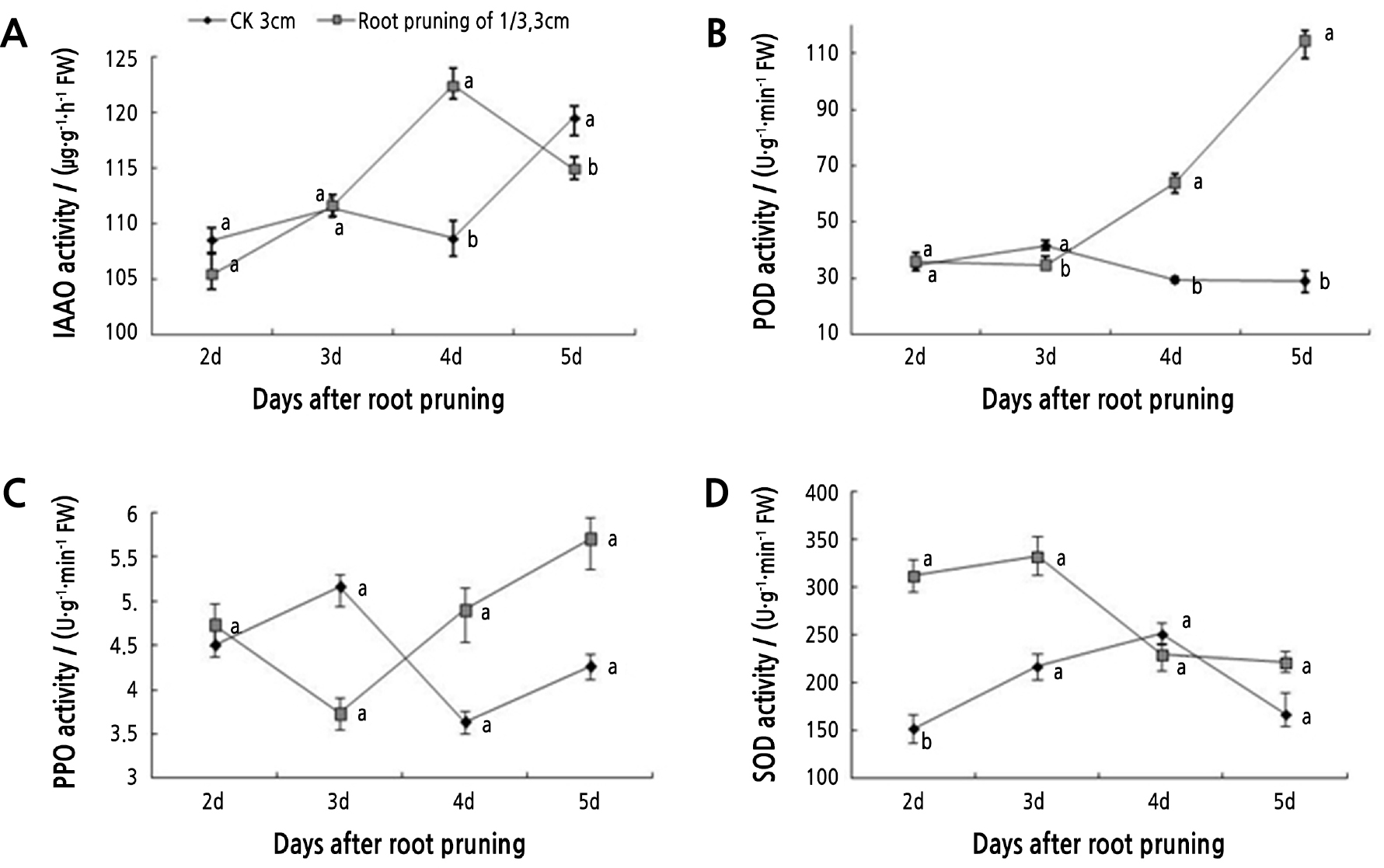
Fig. 4.
Tree peony seedlings with roots 3 cm in length and pruned by 1/3 were analyzed for oxidase enzyme activity. Vertical bars indicate standard error. Each value is the mean obtained from replicates. CK 3 cm indicates the control tree peony seedlings with 3-cm long roots that were unpruned. Values measured on the same day with the same letter are not significantly different according to Duncan’s multiple range test at p < 0.05 (n = 3).
The levels of four plant growth regulator hormones were analyzed over five days after root pruning. IAA levels were significantly higher in pruned roots on day 5, while the level in the control remained basically steady (Fig. 5A). After root primordium formation, the content of IAA decreased and then greatly increased, reaching a maximum on day 5 that was 184.10% higher than that of the control. The levels of GA3 started out higher in pruned roots compared to the control, dipped a little on day 3, then increased by day 5 to a level 136.72% higher than that of the control (Fig. 5B). After root primordium formation, the GA3 content in pruned roots increased rapidly, while the level in the control decreased slowly (Fig. 5B). The ZT content in pruned and control roots showed an overall upward trend during the critical period of rooting (3–5 d). The content of ZT in the pruned roots was lower than that in the control, with a significant difference at 4 d, which was 26.77% lower than that of the control (Fig. 5C). ABA content increased and then decreased in pruned and control roots. However, the ABA content in the pruned roots was lower than that in the control. Additionally, the peak in the ABA content in the pruned roots occurred on day 3, one day earlier than in the control (Fig. 5D).
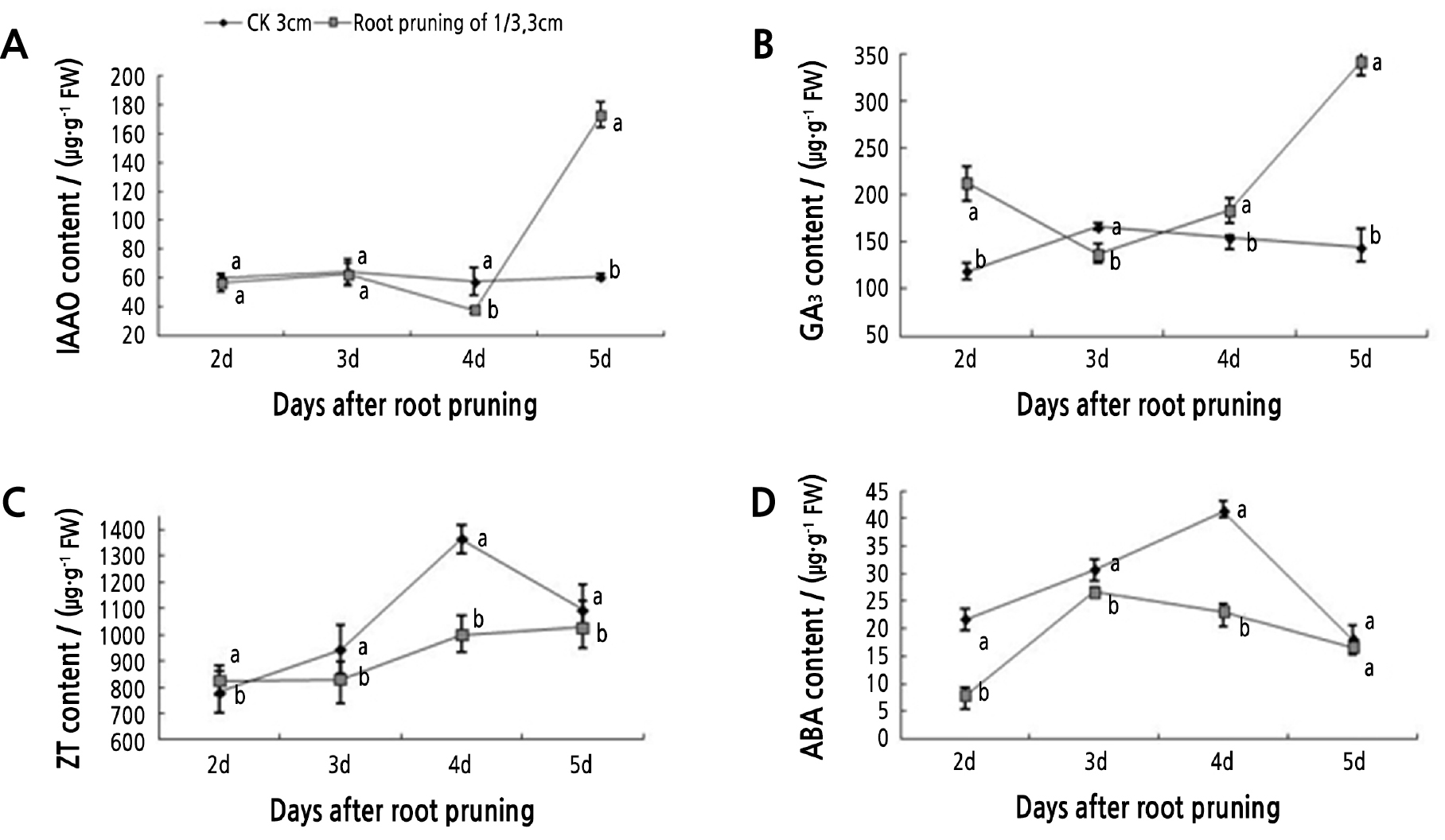
Fig. 5.
Tree peony seedlings with roots 3 cm in length and pruned by 1/3 were analyzed for endogenous hormone content. Vertical bars indicate standard error. Each value is the mean obtained from replicates. CK 3 cm indicates the control tree peony seedlings with 3-cm long roots that were unpruned. Values measured on the same day with the same letter are not significantly different according to Duncan’s multiple range test at p < 0.05 (n = 3).
The IAA/ZT, IAA/GA3, and IAA/ABA ratios were determined in the pruned and unpruned roots (Fig. 6). During the critical rooting period (days 3 to 5), the IAA/ZT ratio of the pruned roots showed a slight decline-then-rise trend and reached the maximum on the 5th day, while it declined in the control. The IAA/GA3 ratio showed a fluctuating upward trend. After the root primordium was formed, the ratio decreased first and then climbed, while the control decreased and then increased to a lesser extent. The IAA/ABA ratio showed consistent trends between the pruned and control roots, although the value was higher for the pruned roots. In the pruned roots, the ratio decreased after the root primordium formed (days 3 to 4) and then increased dramatically to the maximum value on the 5th day.
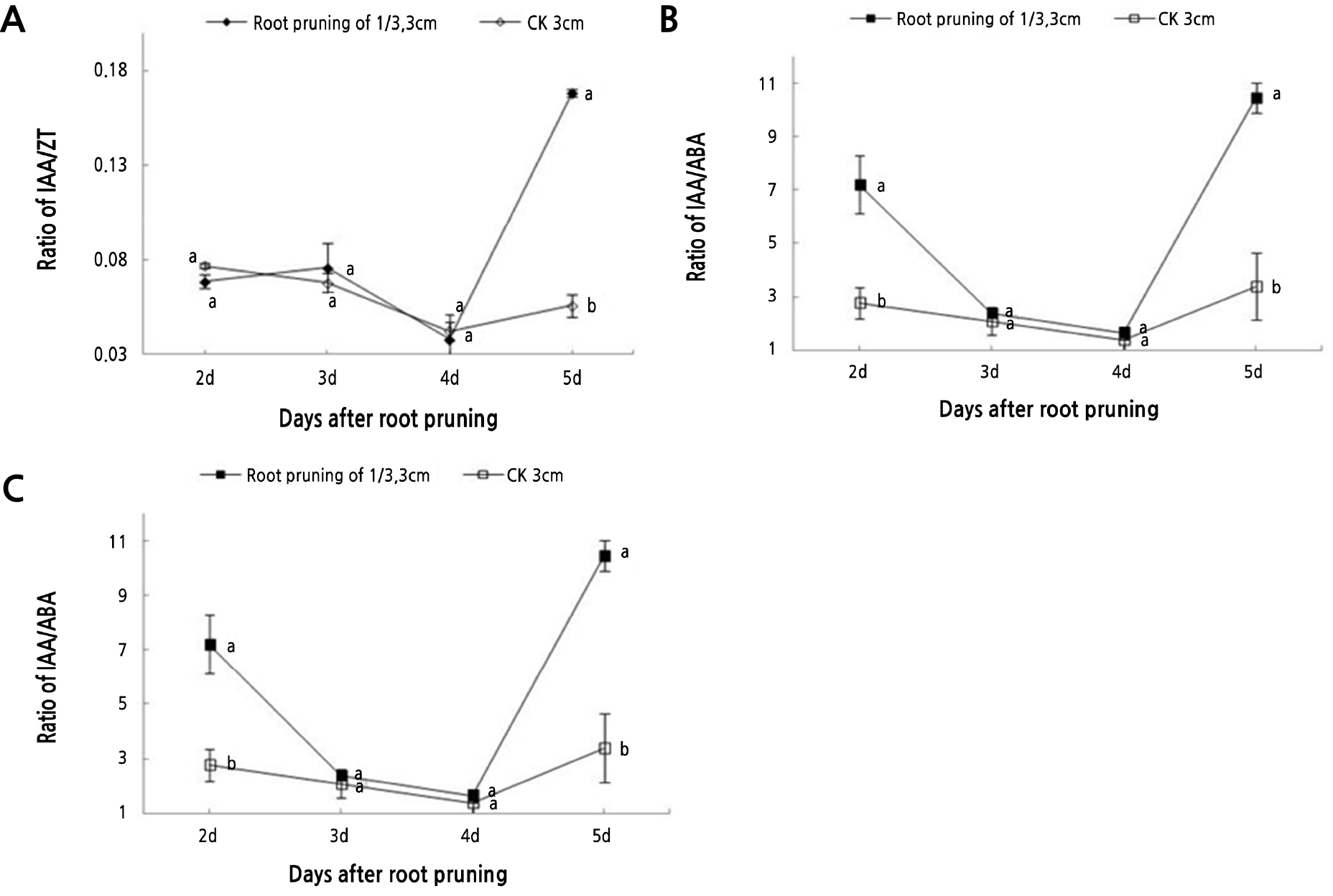
Fig. 6.
The hormone levels in tree peony seedlings with roots 3 cm in length and pruned by 1/3 were used to calculate the IAA/ZT, IAA/GA3, and IAA/ABA ratios. Vertical bars indicate standard error. Each value is the mean obtained from replicates. CK 3 cm indicates the control tree peony seedlings with 3-cm long roots that were unpruned. Values for each day with the same letter are not significantly different according to Duncan’s multiple range test at p < 0.05 (n = 3).
Discussion
In this study, portions of the radicle of the tree peony cultivar ‘Fengdanbai’ were removed to analyze the effects of root pruning on future root architecture. Histological examination of the root structure showed that lateral root formation in peony seedlings occurred mainly 3 to 5 days after root pruning. The root primordia mainly formed on the 3rd day after rooting in the upper 0.5–1.0 cm of the radicle. This coincides with the reported timing of root primordia in tree peony seedlings germinated in vitro in a test tube (He et al., 2011).
In this study, three aspects of root architecture were measured: the number of lateral roots, the root thickness, and the days to lateral root emergence. Four pruning treatments (1/5, 1/3, 1/2, and 3/4) on roots of the same length were compared, with a 1/3 pruning showing the most robust effects. In a second experiment, roots of different lengths (1, 2, 3, or 4 cm) were trimmed by 1/3 their length. The length of the root and the position of the cut differentially affected lateral root growth. The treatment that yielded the most robust peony roots was pruning a 3-cm root by 1 cm, or 1/3 of its length. This treatment yielded the highest number of lateral roots, the greatest rooting rate, and root thickness, indicating that moderate root breaking can promote lateral root initiation and root thickening, which would improve the quality of the root system by increasing the absorption area (Ma et al., 2009).
Respiration activity is an important physiological indicator of plant growth and can be used to detect root vitality. Studies have shown that root pruning can significantly increase root respiratory activity and promote root growth (Li, 2002). Pruning a 3-cm long root radicle by 1/3 increased the respiratory activity higher than in the control roots. The average root respiratory activity during the rooting period was nearly 75% higher than that of the control. These results were consistent with those in tobacco (Nicotiana tabacum L.) showing that root vigor was significantly improved with root cutting (Li et al., 2014).
Studies have shown that changes in POD, PPO, IAAO, SOD, and other related oxidases are closely related to the formation of adventitious roots (Elmongy et al., 2017). POD is a key factor in the formation of root primordia. Gao et al. (2012) found that changes in POD activity were closely related to the rooting of Ceiba pentandra (L.) Gaertn. cuttings and that higher POD activity during adventitious root formation was conducive to rooting. Zhang (2011) found that POD activity of the high-yield poplar tree (Populus tomentosa) significantly increased after root pruning, which indicated that root breaking was closely related to the change of POD activity. In tree peony seedlings, POD activity changed after root pruning. Three to five days after root pruning, POD activity increased, indicating that high levels of POD activity favored adventitious root initiation and elongation. Root pruning increased the POD activity in tree peony seedling roots, which was consistent with POD activity changes in peanut (Arachis hypogaea) roots (Chen et al., 2014). The oxidase PPO catalyzes the reaction of phenolic substances with IAA to form IAA-phenolic acid complexes, which are considered rooting cofactors. In this study, the overall trends in PPO and POD activities were similar 2–5 d after root pruning treatments. The PPO activity prior to the initiation of lateral root primordium was low, and then increased. An increase in PPO activity during the rooting process of tree peony tissue is conducive to the formation of adventitious roots (He et al., 2011), indicating that increased PPO activity could be used as an indicator of adventitious root formation. After root pruning, the SOD activity of tree peony seedlings peaked on the 3rd day during root primordium induction, indicating that high SOD activity could promote adventitious roots, which was consistent with results on Cyclocarya paliurus L. and Lonicera japonica L., in which SOD activity was positively correlated with the rooting rate (Zhang, 2009; Li et al., 2015).
IAAO activity regulates IAA levels in plants, and its effect on rooting would depend on IAA levels. IAA is the main hormone that promotes the formation of adventitious roots. In this study, IAAO activity increased in tree peony seedling roots that were trimmed by 1/3 when they were 3 cm long. IAAO activity increased during root primordium induction, then decreased during the elongation phase. We presumed that some IAA may be degraded by IAAO, which then allows the formation of root primordium, although a return to a higher level of IAA would benefit root elongation. In pruned tree peony seedling roots, the content of IAA increased during root primordium induction, decreased after root primordium formation, and then reached its maximum, which was 184% higher than that of the control, when the adventitious roots broke through the epidermis (on day 5). This phenomenon between IAAO activity and IAA levels was also found in hazelnut (Corylus heterophylla Fisch.) (Hu et al., 2008), Populus tomentosa Carrière (Song et al., 2001), and Betula platyphylla Suk (Zhan et al., 2001).
The dynamic changes in endogenous hormone content during rooting are closely related to the formation of adventitious roots. GA3 can accelerate stem elongation and promote plant growth and maturation. In this study, the GA3 content of tree peony seedling roots, pruned when under 3 cm long by 1/3, was opposite to that of the unpruned control. GA3 activity decreased during root primordium induction, indicating that high levels of GA3 may inhibit the formation of root primordium, which was consistent with findings in Picea abies (L.) Karst. (OuYang et al., 2015) and Sabina davurica (Pall.) Ant. (Guo et al., 2012). After the formation of root primordium, the content of GA3 increased slightly. It could be speculated that a high level of GA3 promoted the initiation and elongation of adventitious roots and then regulated the occurrence of adventitious roots. On the other hand, numerous studies have shown that zeatin (ZT) mainly inhibits the differentiation and formation of root primordia (Guo et al., 2012; Feng et al., 2012), but other studies have shown that higher ZT promotes adventitious root formation. In peony seedling roots pruned by 1/3 when they were 3 cm long, the ZT content was overall lower than in the unpruned roots and showed an upward trend after lateral root initiation. This might be because the cytokinin level decreased after root pruning when the root tip, the main source of cytokinins, was removed. After the wound healed, the root system started to produce new cytokinin regulated by its own mechanism. In the early stage of root primordium induction, low levels of ZT was conducive to the formation of root primordium. The content of ZT increased during the critical period of adventitious root formation (3–5 days after root pruning), possibly promoting the formation and elongation of adventitious roots. Therefore, the effect of ZT was different during the adventitious rooting phase and the elongation phase. This conclusion is similar to that of Wang (1992). As an important growth inhibitor in plants, ABA is closely related to the formation of adventitious roots and mainly inhibits cell division and elongation (Yang and Pan, 2015; Cao, 2015). In tree peony seedling roots pruned by 1/3 when they were under 3 cm, the ABA content increased before root primordium formation. This may be due to the large amount of ABA produced for root wound suberization, similar to results in Kiwi (Actinidia deliciosa) fruit (Han et al., 2018). After root primordium formation, ABA levels declined, indicating that new roots could reduce ABA content and facilitate adventitious root formation.
Growth regulators are not present alone in plants, and they regulate the development of plants by mutual promotion or antagonism (He et al., 2002; Wu et al., 2006; Steffens et al., 2006). IAA plays a key role in the process of adventitious root formation. Tracking changes in the ratios of IAA/GA3, IAA/ZT, and IAA/ABA can better reflect the balance between endogenous hormones during adventitious root formation (Xie et al., 2009; Yan et al., 2010; Zhang et al., 2014). Recent studies reported that IAA content, the IAA/ABA ratio, and the IAA/GA3 ratio were better indicators of adventitious root formation in Thuja occidentalis Linn., Castanea mollissima cv. ‘Yanshanhong’, and C. paliurus (Liu et al., 2010; Hou et al., 2010; Li et al., 2015). In tree peony seedling roots pruned by 1/3 when they were 3 cm long, the IAA/GA3 and IAA/ZT ratios and the IAA content during the root primordium induction period were all increased gradually and similarly. Meanwhile, the IAA/ABA ratio decreased gradually during the same period, which may be due to the transient increase in ABA caused by the root pruning wound. The change in the IAA/ABA ratio after root primordium formation was consistent with the changes in IAA, that was, it rose rapidly after a decline, during which ABA gradually decreased. IAA accumulated continuously and played a leading role in the occurrence of adventitious roots. Xie et al. (2009) reported that a higher IAA/GA3 ratio contributes to the induction and elongation of adventitious roots in C. paliurus and that GA3 was not a key factor in this process. In this study, the change in the IAA/GA3 ratio following pruning showed a trend of rising-decreasing-rising during the process of adventitious roots. The trends for IAA and GA3 were different during the formation of adventitious roots. Therefore, the IAA/GA3 ratio changed more than the content of IAA or GA3, and this ratio could better reflect the changes in endogenous hormones during the process of adventitious rooting. It is clear that the higher IAA/GA3 ratio contributed to the induction and elongation of adventitious roots. After root primordium formation, the trends of the IAA/ZT ratio and IAA content were similar, both decline-rise trends. The IAA/ZT ratio increased during adventitious root elongation, which may be due to the rapid increase in IAA content after the formation of new root tips.
In summary, this study analyzed the effects of different root pruning treatments on lateral root formation of tree peony seedlings and on root vitality, enzyme activity, and endogenous hormone levels during adventitious root formation. The results showed that pruning tree peony seedling roots when they were 3 cm in length by removing 1/3 of their overall length increased the number of lateral roots, the degree of root thickening, and root vitality as measured by respiration activity. Removing the end of the root tip significantly altered the levels of endogenous hormones and the activities of several oxidases during rooting. Specifically, there were decreases in SOD activity and ABA and ZT levels, and significant increases in the activities of the oxidases POD, PPO, and IAAO and in the contents of IAA and GA3. The ability to propagate the tree peony cultivar ‘Fengdanbai’ from seed is limited due to lateral root dysplasia. This experiment represents an important step in being able to germinate seeds and transplant seedlings of ‘Fengdanbai’, although the effects of an early root pruning treatment on later root and plant growth and development still need further study.
