

서 언
재료 및 방법
실험재료 및 처리 환경
생육조사
상토 및 식물체 무기원소 분석
통계처리
결과 및 고찰
파종 39일 후 생장
파종 70일 후 생육
상토의 EC 변화
파종 70일 후 잎의 무기성분 함량
서 언
고추(Capsicum annuum L.)는 풋고추 및 조미채소용 건고추 생산을 목적으로 국내에서 재배되며, 풋고추는 시설재배로 건고추는 노지재배로 생산된다. 2023년 기준 건고추 생산을 위한 재배면적은 풋고추보다 57배 높은 27,132ha이었다(KOSIS 2025). 노지재배 방법으로 건고추 생산을 위한 홍고추를 재배할 경우 전문 육묘장에서 첫 꽃이 필 시기까지 육묘한 묘를 농가에 보급하고, 무상기간에 노지에 정식한 후 본포 재배를 한다.
연속 개화 및 착과가 이루어지는 고추의 생리적 특성상 육묘 후 본포에 정식함으로써 노지에 직파하는 경우 보다 재배기간 연장을 통한 수확량 증가가 가능하다. 국내 전문 육묘장들은 주로 2월에 고추 종자를 파종한 후 약 80일 정도 육묘하고(Kim et al. 2015; Lee et al. 2018) 농가에 묘를 공급하지만 이 시기의 기온이 낮은 이유로 가온하여 육묘환경을 조절하여야 한다. 그러나 육묘장 별로 환경 조절 방법이 다르고 주·야간 설정 온도가 달라 고추 묘의 생장속도 및 생장량이 다르다. 또한 육묘기간에 생장을 촉진시켜 본포 정식에 적합한 크기에 도달하는 기간이 짧아질 경우 난방비, 노동비 및 기타 묘 생산을 위한 투입 비용이 절감되어 생산비가 감소하고 육묘 농가의 수익이 증가한다.
육묘기간의 광도(Zhang et al. 2003), 관수 수준(Jeong et al. 2020; Moon et al. 2023), 상토(Choi et al. 1997), 비료 종류(Park et al. 2025b), 공급 양액의 EC 수준(Park et al. 2017; Park et al. 2025a), 그리고 육묘기간(Kim et al. 2015)이 고추 묘 생장에 영향을 미친다. 기온이 묘 생장에 미치는 영향과 관련하여서는 육묘기간 중의 초장을 조절하기 위한 수단으로 주·야간 온도차(Difference between day and night temperatures, DIF)를 위주로 한 연구가 다수 이루어졌다(Stavang et al. 2005; Kim et al. 2013; Yang et al. 2016; Ohtaka et al. 2020; Song et al. 2023).
그러나 고추 육묘를 위한 대부분의 국내 육묘장들은 주·야간 온도차를 인위적으로 완벽하게 조절할 수 있을 만큼 시설환경 자동화 시설이 잘 갖추어지지 않을 뿐만 아니라 주·야간 온도차를 조절하기 위한 과도한 난방비 투입 역시 문제가 될 수 있다. 또한 적극적으로 시설환경을 조절할 수 있어 육묘온도를 높게 설정하는 것이 가능한 농가 또한 난방비 지출을 고려하여 설정온도를 낮게 설정한 상태로 육묘하는 농가가 많다. 그러므로 시설환경이 자연상태의 대기환경에 큰 영향을 받아 주간온도는 높고 야간온도는 낮게 유지된다. 한편 주·야간 온도차가 8°C 이상 급격하게 높아지면 지하부의 생장에 비해 지상부의 도장의 우려가 크고(Lim et al. 1996), 작물 스트레스로 인해 수박의 묘소질이 불량해졌으며, 적정 주·야간 온도는 각각 25°C와 20°C인 +5DIF였다고 보고하였다(Kwack and An 2021). 대부분의 농가들은 대기온도 조건이 고추 묘 생장에 큰 영향을 미침을 알고 있지만 정밀하게 수행하여 보고된 연구결과가 없고, 농가에서 참고할 자료가 없는 상황이다.
따라서 주간온도와 야간온도의 차이를 5°C로 유지하면서 저온(주간/야간, 15/10°C), 중온(주간/야간, 20/15°C) 및 고온(주간/야간, 25/20°C)으로 온도를 조절하여 홍고추 3 품종을 육묘하면서 묘 생장과 무기원소 함량에 미치는 영향을 구명하고 고추 육묘를 위한 기초 자료를 제시하기 위하여 본 연구를 수행하였다.
재료 및 방법
실험재료 및 처리 환경
국내에서 육성되고 건고추 생산용인 3 종류의 홍고추, ‘AT Sinhotan’(Asia Seed Co., Ltd, Seoul, Korea), ‘Allbokhap’(Dana Co., Ltd, Seoul, Korea) 및 ‘Callatan’(Sakata Korea Co., Ltd, Seoul, Korea)을 실험대상으로 삼았다. 한아름B 상토(Shinsung Mineral Co., Ltd, Seongnam, Korea)를 72셀 트레이에 충진하고, 종자를 파종하였으며, 파종 후 버미큘라이트(Green Fire Chemials Co., Ltd, Hongsung, Korea)로 2mm 정도 복토하였다. 이후 28°C와 암상태로 조절한 생장상(HB-301L, Hanbaek Scientific Technology Co., Bucheon, Korea)에 1주일간 두었다. 발아 후 광합성유효광량자속이 300µmol·m-2·s-1 전후, 일장을 16시간으로 조절한 생장상에서 육묘하였다.
온도의 경우 주·야간 온도차를 +5DIF로 동일하게 유지하면서 주간(day, D)/야간(night, N) 온도를 변화시켜 각각 15°C(D)/10°C(N), 20°C(D)/15°C(N) 및 25°C(D)/20°C(N)로 조절한 3처리로 실험하였다. 양액은 18-18-18(N-P2O5-K2O, Haifa Chemicals Ltd, Matam-Haifa, Israel) 비료로 조제하였으며, 질소 기준 150mg·L-1로 농도를 조절한 양액을 공급하였다. 이 때 별도로 미량원소복합제(MgO 3%, 킬레이트 Fe 5%, Zn 2.5%, Mn 2.5%, Cu 0.5%, B 1%, Mo 0.05%, Nutrichem Kombi-F, Grobbendonk, Belgium)를 0.2g·L-1 첨가하였다. 양액의 pH와 EC는 용해 후 각각 6.21과 1.24dS·m-1였지만 질산으로 pH를 5.9로 조정하여 처리하였고, 이 양액의 EC는 1.24dS·m-1였다. 파종 후 11일째부터 상기의 양액을 주 1회, 칼슘을 보충하기 위해 파종 후 7주째부터 Proplex Ca 20(Plant Health Ltd, Nottingham, United Kingdom)을 1/1,500로 희석(pH 5.7, EC 0.92dS·m-1)하여 상기의 양액과 겹치지 않게 주 1회 각각 두상관비하였다.
생육조사
72셀 플러그 트레이에 파종한 후 70일간 육묘하였으며 파종 후 39일과 70일째 2회 생육을 조사하였다. 조사항목은 식물체의 초장, 경경, 엽수, 엽면적, 엽색, 엽록소 함량, 지상부의 생체중 및 건물중 등이었다.
생육조사에서 초장은 상토 표면에서 생장점까지의 길이를 측정하였으며, 경경은 자엽의 약 1cm 하단의 굵은 부위를 버니어캘리퍼스(CD-15APX, Mitutoyo Corp., Kanagawa, Japan)를 사용하여 측정하였다. 엽수는 전개된 모든 본엽만 세었고, 전개엽 중 가장 큰 잎의 엽록소 함량을 SPAD meter(SPAD-502, Minolta, Osaka, Japan)로 측정하였다. 엽면적은 개체 별 모든 잎을 분리하여 엽면적 측정계(LI-3100C, LI-COR Inc., NE, USA)를 사용하여 측정한 후 합산하였다. 엽색은 색도색차계(Chroma Meter CR-400, Konica Minolta Inc., Tokyo, Japan)를 사용하여 Kang et al.(2005)의 방법에 따라 측정하였다. 개화수는 각 식물체에 착생된 꽃 중 개화된 꽃만 계수하였으며 화뢰는 제외시켰다. 식물체의 무게는 저울(EL3002-IC, Mettler Toledo, Langacher, Swiss)로 생체중을 측정한 다음 75°C 건조기(HB-502L, Han Baek Scientific Co., Bucheon, Korea)에서 24시간 건조시켜 건물중을 측정하였다. 묘소질의 정도는 Dickson et al.(1960)에 따라 아래 식에 따라 Dickson quality index(DQI) 값으로 구하였다. DQI = TDW/(PH/SD + SDW/RDW) [TDW(total dry weight); 총 건물중(g), PH(plant height); 초장(cm), SD(stem diameter); 경경(mm), SDW(shoot dry weight); 지상부 건물중(g), RDW(root dry weight); 지하부 건물중(g)]. 처리간 생육 정도는 파종 후 70일째 사진을 촬영하였다.
상토 및 식물체 무기원소 분석
상토의 전기전도도(electrical conductivity, EC)와 pH는 파종 후 39일과 70일째에 표토를 제거하고 상토를 수집하여 풍건시켰다. 풍건 상토와 증류수를 1:10(w/w)의 비율로 혼합하고 2시간을 기다렸다가 추출한 후 측정(multimeter CP-500L, Istek Inc., Seoul, Korea)하였으며 Park et al.(2017)의 방법에 따라 수행하였다.
식물체 무기원소 분석은 파종 후 70일째 생육조사 후 건조시킨 잎을 분쇄하여 시료로 사용하였다. 분쇄한 시료의 전질소(T-N) 함량은 Kjeldahl 법(Eastin 1978)으로 분석하였다. 한편 500°C 전기로(WiseTherm FSC-22, Daihan Scientific Co., Ltd, Wonju, Korea)에서 6시간 건식회화시킨 시료를 0.5N HCl 용액으로 포집하여 질소를 제외한 무기원소 분석에 사용하였다. K, Ca, Mg, Cu, Fe, Mn 및 Zn 함량을 원자흡광분광광도계(AA-7000, Atomic Absorption Spectrophotometer, Shimazu Corp., Kyoto, Japan)로, P 함량을 분광광도계(UV mini-1240, Shimadzu Corp., Kyoto, Japan)를 사용하여 분석(Chapman and Pratt 1961)하였으며, 분석과정은 Park et al.(2020)의 방법에 준하였다.
통계처리
온도 처리에 따라 72셀 플러그 트레이를 품종별로 각각 3개씩 완전임의배치 하였으며, 처리당 3 트레이에서 각각 5개체씩 3반복으로 선발하여, 총 15주의 생장을 조사하였다. 온도 처리와 품종 간 비교는 SPSS Statistics(Ver. 29.0.2.0, IBM, Armonk, NY, USA)을 이용하여 분산분석(analysis of variance, ANOVA)하였고, 처리 간 평균은 Costat 프로그램(Ver. 6.311, CoHort Software, Monterey, CA, USA)을 이용하여 Duncan의 다중검정법으로 p < 0.05 수준에서 비교하였다.
결과 및 고찰
파종 39일 후 생장
파종 39일 후 다양한 온도 처리가 플러그 묘 생장에 미치는 영향을 조사한 결과 주·야간 온도를15°C(D)/10°C(N)로 조절한 처리구에서 초장이 가장 짧았고 품종에 관계없이 4cm 미만으로 조사되었다(Fig. 1A). 그러나 주·야간 온도가 높아질수록 초장이 길어졌으며 25°C(D)/20°C(N) 처리구의 ‘AT Sinhotan’ 고추가 12.2cm로 가장 길었고, 동일한 온도 처리구 내에서 ‘AT Sinhotan’ 고추가 다른 두 품종보다 초장이 유의하게 길었다. 세 품종의 줄기 굵기(경경)은 주·야간 온도를 20°C(D)/15°C(N) 처리구에서 가장 굵었고 25°C(D)/20°C(N), 15°C(D)/10°C(N) 처리구 순으로 가늘어졌다(Fig. 1B). 20°C(D)/15°C(N)와 25°C(D)/20°C(N) 처리구 내의 경경은 품종 간 유의미한 차이가 없었지만 15°C(D)/10°C(N)로 온도를 조절한 경우 ‘Callatan’의 경경이 유의하게 가늘었고, 이는 저온에 대한 품종간 내성 차이가 원인이라고 생각한다. 또한 신장 생장과 비례하여 마디수가 많아졌으며, 25°C(D)/20°C(N) 처리구에서 5.6±0.2개였다(Fig. 1C). 주·야간 온도가 높아질수록 지상부 생체중이 무거웠으며, 초장이 가장 길었던 ‘AT Sinhotan’이 가장 무거웠고 동일한 처리구[25°C(D)/20°C(N)] 내의 ‘Callatan’이 통계적으로 유의하게 가벼웠다(Fig. 1D). 그러나 지상부 건물중은 생체중과 동일한 경향을 나타내지 않았다. 즉, 20°C(D)/15°C(N) 처리구의 ‘AT Sinhotan’이 식물체당 0.19g으로 산술적으로는 가장 무거웠지만 20°C(D)/15°C(N)와 25°C(D)/20°C(N) 처리구의 모든 품종 간에는 통계적으로 유의한 차이가 없었다(Fig. 1E). 고추 플러그 묘의 파종 39일 후 DQI는 온도 처리구간 차이가 없었다. 그러나 이는 온도 처리구 간 건물중의 차이가 크지 않았던데 비해 15°C(D)/10°C(N) 처리구의 초장이 20°C(D)/15°C(N)와 25°C(D)/20°C(N) 처리구의 초장보다 각각 54–60%와 68–72% 작았던 이유로 DQI가 높게 나타났다고 판단한다(Fig. 1F).
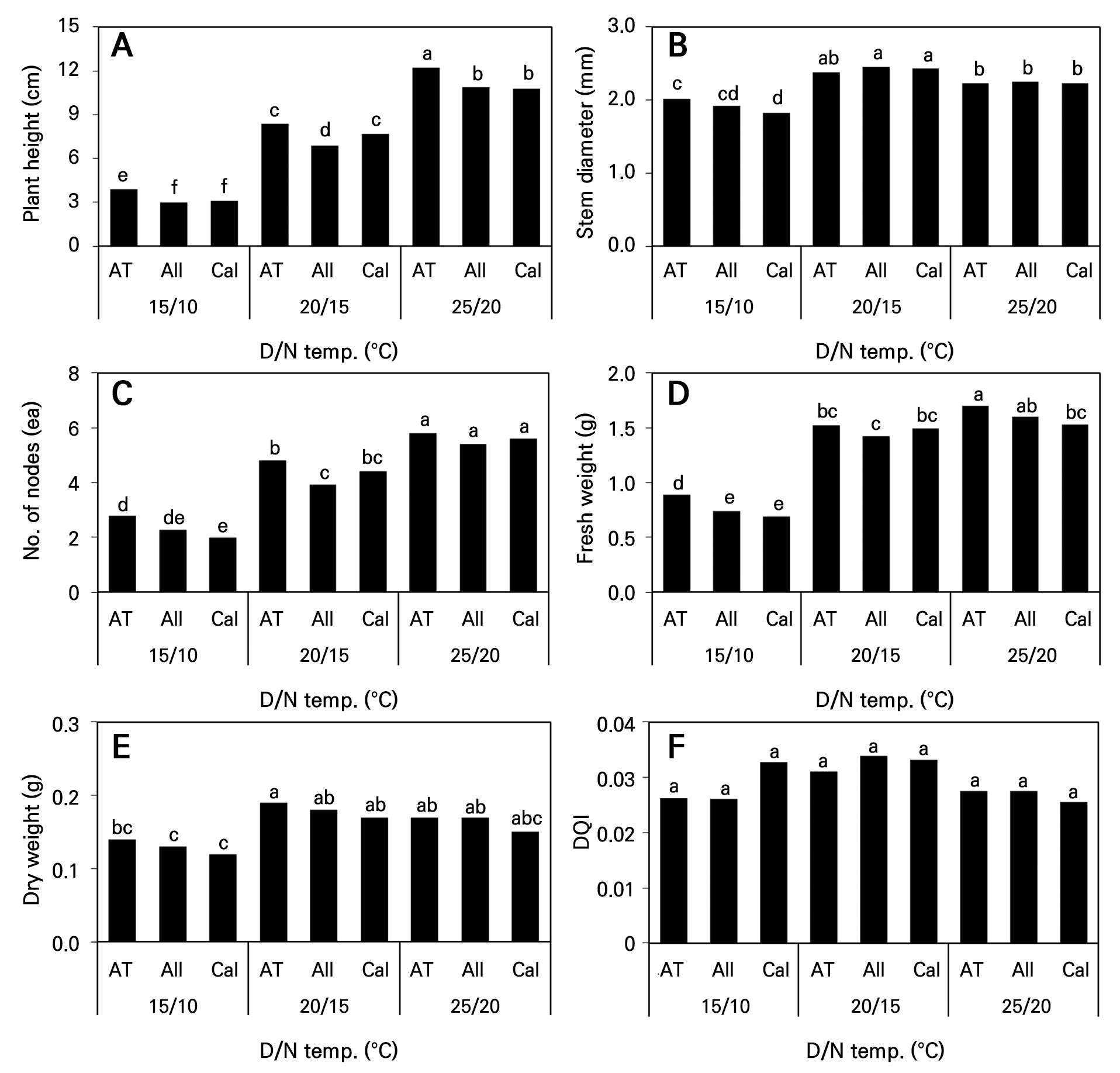
Fig. 1.
Growth indices of three hot pepper cultivars as influenced by day and night temperature changes at 39 days after sowing. Different letters within columns indicate significant difference based on Duncan’s multiple range test, p < 0.05 (n = 3). AT, ‘AT Sinhotan’; All, ‘Allbokhap’; Cal, ‘Callatan’.
파종 후 39일의 육묘기간 중 발생된 엽수는 주·야간 온도 25°C(D)/20°C(N) 처리구의 ‘AT Sinhotan’ 고추가 7.3개로 가장 많았지만 20°C(D)/15°C(N) 처리구의 ‘AT Sinhotan’ 및 ‘Callatan’의 엽수와는 통계적으로 유의한 차이가 없었다(Fig. 2A). 처리 온도가 높아질수록 식물체 당 발생한 모든 잎의 엽면적이 넓어졌지만 25°C(D)/20°C(N) 처리구에서는 품종 간 차이가 뚜렷하지 않았다(Fig. 2B). SPAD 값은 엽면적이 가장 작았던 15°C(D)/10°C(N) 처리구에서 가장 컸으며 온도가 높아질수록 작아지는 경향이었다(Fig. 2C). Hunter L* 값은 ‘AT Sinhotan’ 고추의 주간 온도 20°C 이상에서 측정값이 높아 다른 두 품종보다 엽색이 밝았다(Fig. 2D).
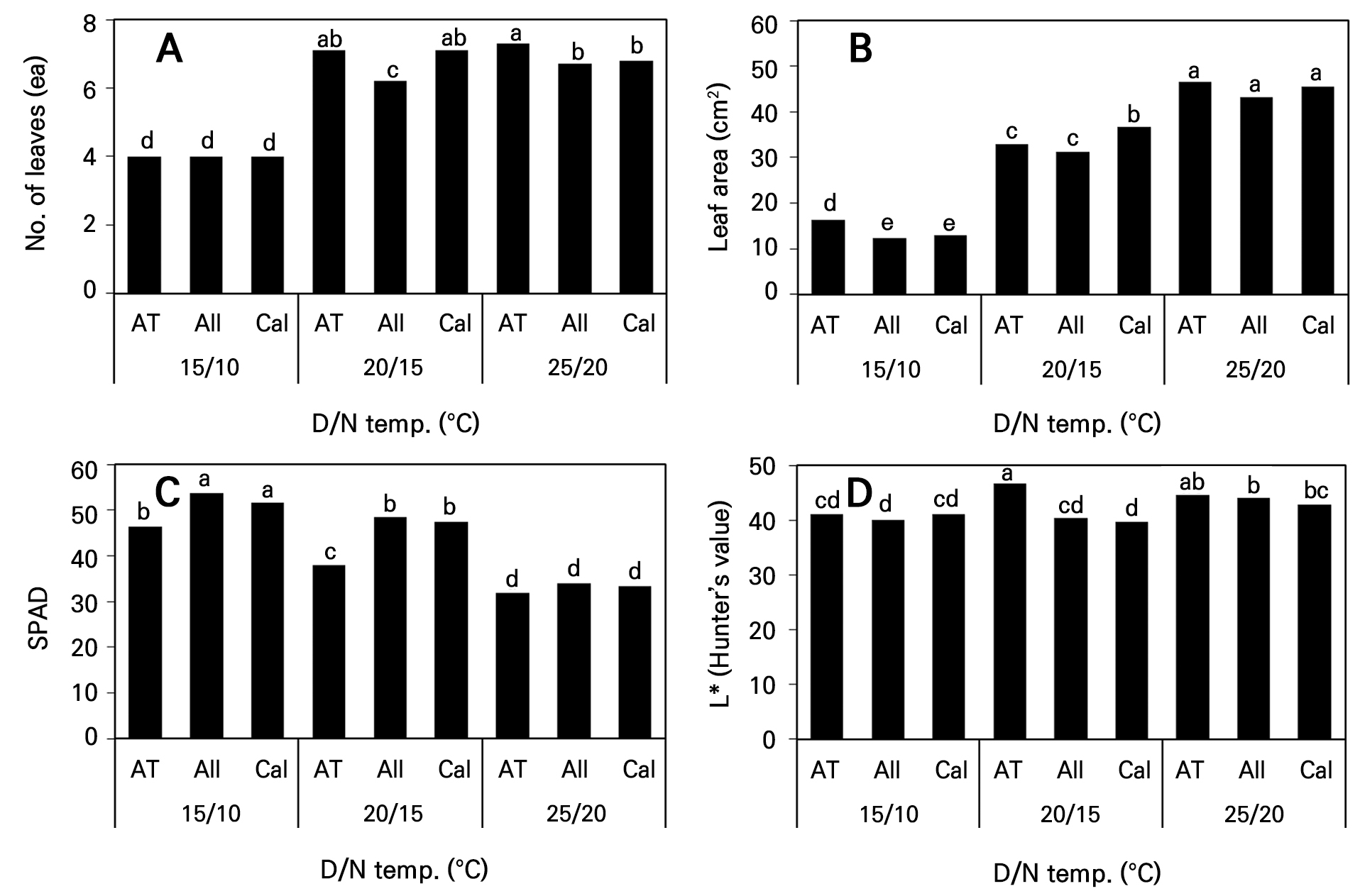
Fig. 2.
Leaf number, leaf area, SPAD, and Hunter’s L* of three hot pepper cultivars according to day and night temperature changes at 39 days after sowing. Different letters within columns indicate significant difference based on Duncan’s multiple range test, p < 0.05 (n = 3). AT, ‘AT Sinhotan’; All, ‘Allbokhap’; Cal, ‘Callatan’.
파종 70일 후 생육
파종 후 70일 동안의 육묘기간 중 주·야간 온도가 높을수록 초장이 길어졌고, 25°C(D)/20°C(N) 처리구의 ‘AT Sinhotan’ 고추가 29.4cm로 가장 길었으며, 15°C(D)/10°C(N) 처리구는 상대적 저온으로 생장이 억제되어 3 품종의 초장이 약 5cm로 조사되었다(Fig. 3A). 줄기 굵기는 20°C(D)/15°C(N) 처리구에서 다른 온도 처리구 보다 세 품종 모두 유의하게 굵었으며, 세 품종 간에는 ‘Callatan’ 고추가 3.57mm로 가장 굵었다(Fig. 3B). 그리고 15°C(D)/10°C(N)와 25°C(D)/20°C(N) 처리구는 모든 품종에서 줄기 굵기가 약 2.8mm로 품종간 차이가 뚜렷하지 않았다. 마디수는 20°C(D)/15°C(N)와 25°C(D)/20°C(N) 처리구에서 ‘AT Sinhotan’ 고추가 약 11개로 가장 많았다(Fig. 3C). 지상부 생체중은 주·야간 온도가 높을수록 무거웠으나(Fig. 3D) 건물중은 20°C(D)/15°C(N)와 25°C(D)/20°C(N) 처리구 간 유의한 차이가 인정되지 않았고, 20°C(D)/15°C(N)에 비해 25°C(D)/20°C(N) 처리구에서 식물체 내 함수율이 상대적으로 높았음을 알 수 있었다(Fig. 3E). 우량묘를 판단하는 주요 지표 중 하나인 DQI는 15°C(D)/10°C(N) 처리구에서 가장 컸고 20°C(D)/15°C(N) 및 25°C(D)/20°C(N) 순으로 낮아졌다(Fig. 3F). 그러나 상대적으로 낮은 온도인 15°C(D)/10°C(N) 처리구는 3 품종의 지하부 생장에 비해 상대적으로 지상부 생장(초장 6cm 이하)이 지나치게 억제되어, 이로 인해 가장 높게 계산되었다고 판단하였으며, 본 실험의 DIF 처리 시 우량묘를 판단하기 위한 기준으로 삼기에 다소 미흡한 점이 있다고 생각한다. 오히려 20°C(D)/15°C(N) 처리구의 플러그 묘가 우량묘로서 적합하다고 판단한다. 이는 Lim et al.(1996)이 ‘녹광’ 고추 플러그 육묘에서 주·야간 온도가 각각 22.5°C와 15°C에서 강건한 묘를 생산할 수 있었다고 하여 본 연구와 야간온도는 동일하였으나 주간온도는 2.5°C 더 높았다. 이는 품종별 차이 보다는 실험구 설정이 본 실험과 다소 달랐기 때문이라 판단되었다.
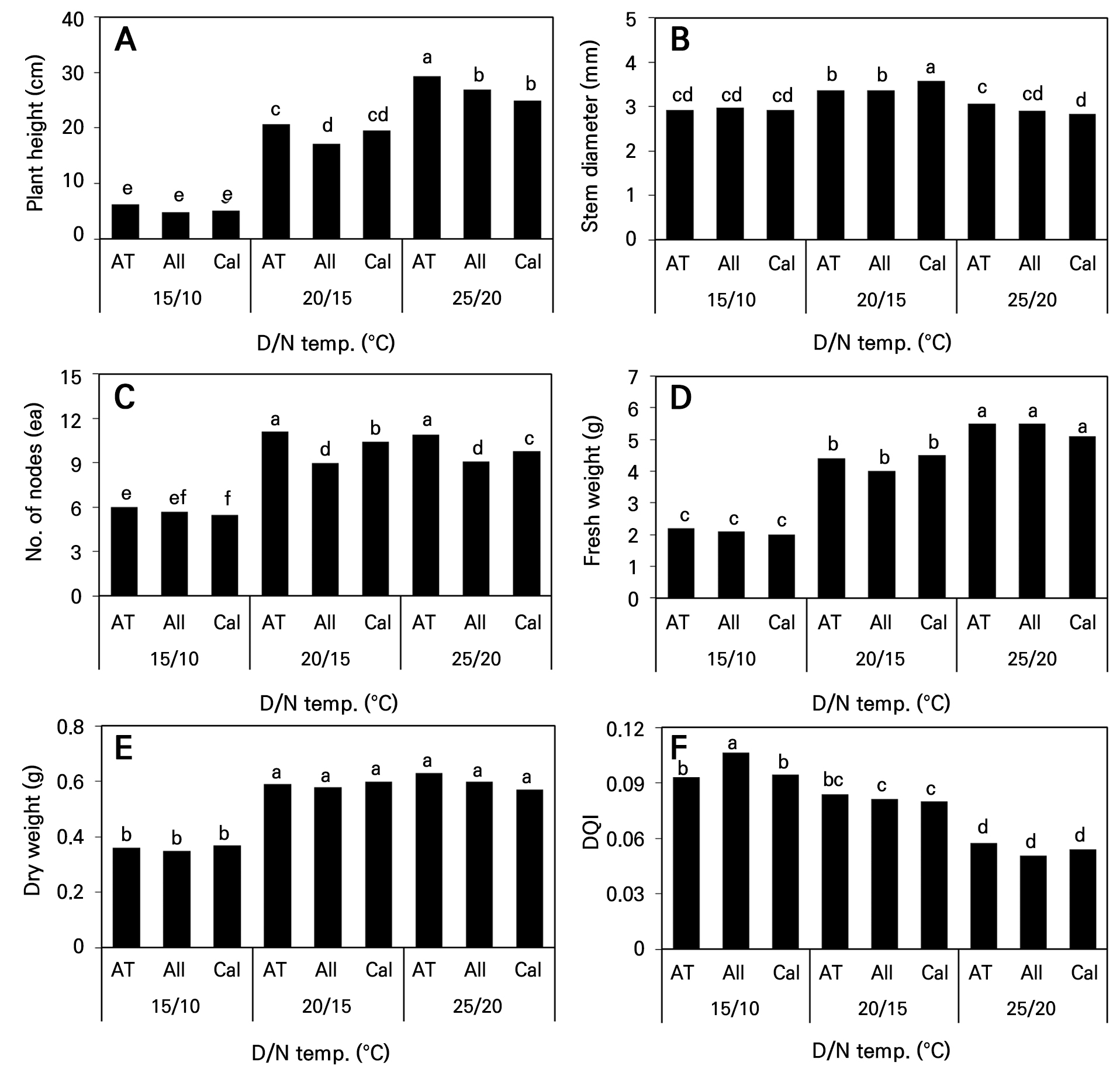
Fig. 3.
Growth indices of three hot pepper cultivars as influenced by day and night temperature changes at 70 days after sowing. Different letters within columns indicate significant difference based on Duncan’s multiple range test, p < 0.05 (n = 3). AT, ‘AT Sinhotan’; All, ‘Allbokhap’; Cal, ‘Callatan’.
육묘 종료 시 조사한 고추의 엽수는 주·야간 온도가 높았던 25°C(D)/20°C(N) 처리구에서 많았으며, ‘Callatan’ 고추의 엽수가 15.8개로 가장 많았고 ‘AT Sinhotan’ 고추와는 통계적으로 유의한 차이가 인정되었다(Fig. 4A). 그러나 파프리카와 오이의 경우 주·야간 온도차 보다 일평균기온이 엽수에 더 큰 영향을 미쳤으며 온도가 16°C에서 약 21°C로 높아짐에 따라 육묘기간 중 묘의 엽수가 증가하였다고 보고한 바 있고(Bakker and van Uffelen 1988; Grimstad and Frimanslund 1993) 본 연구결과에서도 주·야간 온도가 높아질수록, 즉 일평균 온도가 높을수록 엽수는 증가하여 유사한 결과였다. 엽면적은 25°C(D)/20°C(N) 처리구에서 ‘Allbokhap’ 고추가 가장 넓었으며 ‘AT Sinhotan’ 및 ‘Callatan’ 고추 순으로 적어졌다(Fig. 4B). 한편 ‘Callatan’ 고추의 경우 엽수가 많았음에도 불구하고 엽면적이 적었으며 다른 품종에 비해 개별 잎의 크기가 작았음을 의미한다고 판단한다. Bakker and van Uffelen(1988)은 주·야간 온도차 뿐만 아니라 일평균 기온도 파프리카의 엽면적에 영향을 미친다고 보고하였고, Lim et al.(1996)은 일평균 온도가 증가함에 따라 고추의 엽장과 엽폭이 커진다고 보고하여 본 연구에서 온도가 높아질수록 엽면적이 커진 것과 유사한 보고를 한 바 있다. 또한 Kim et al.(2015)은 홍고추 ‘배로따’의 육묘 기간이 45일에서 85일로 길어질수록 엽면적이 2배 이상 증가하였다고 보고하였다. 본 연구에서도 육묘 기간이 파종 후 39일에서 70일로 길어질수록 엽면적이 2–3배 증가하여 유사한 결과를 나타내었다고 판단하였다.
간접적으로 엽록소 함량을 측정하는 SPAD 값은 온도가 낮을수록 높아지는 경향이었고, 생장이 가장 저조했던 15°C(D)/10°C(N) 처리구의 ‘Allbokhap’이 가장 높았으며 동일 온도 처리구 내의 ‘Callatan’과는 유의한 차이가 없었다(Fig. 4C). Lim et al.(1996)은 ‘녹광’ 고추 플러그 육묘에서 주간온도에 관계없이 야간온도가 낮은 처리구에서 엽록소 함량이 높았고, Yang et al.(2021)도 토마토 재배 시 야간온도를 15°C 보다 10°C로 관리하였을 때 엽록소 함량이 더 증가하였다고 보고하였으며 본 연구결과를 뒷받침하였다. 이는 야간온도를 낮춤으로써 호흡량이 감소하여 주간의 광합성으로 생성된 탄수화물의 소모량을 줄여 엽록소 분해를 지연시키거나 축적을 유도하리라 판단된다. 한편 모든 온도 처리구 내에서 ‘AT Sinhotan’이 다른 품종에 비해 SPAD 값이 낮았으며, 온도에 대한 품종 별 생장 반응이 다른 것이 원인이라고 판단하였다. 동일한 품종 내에서 온도가 낮았던 처리가 높았던 처리들 보다 엽색의 명도를 나타내는 Hunter L* 값이 높았으며, SPAD 값이 낮았던 ‘AT Sinhotan’이 다른 품종들 보다 높았다(Fig. 4D).
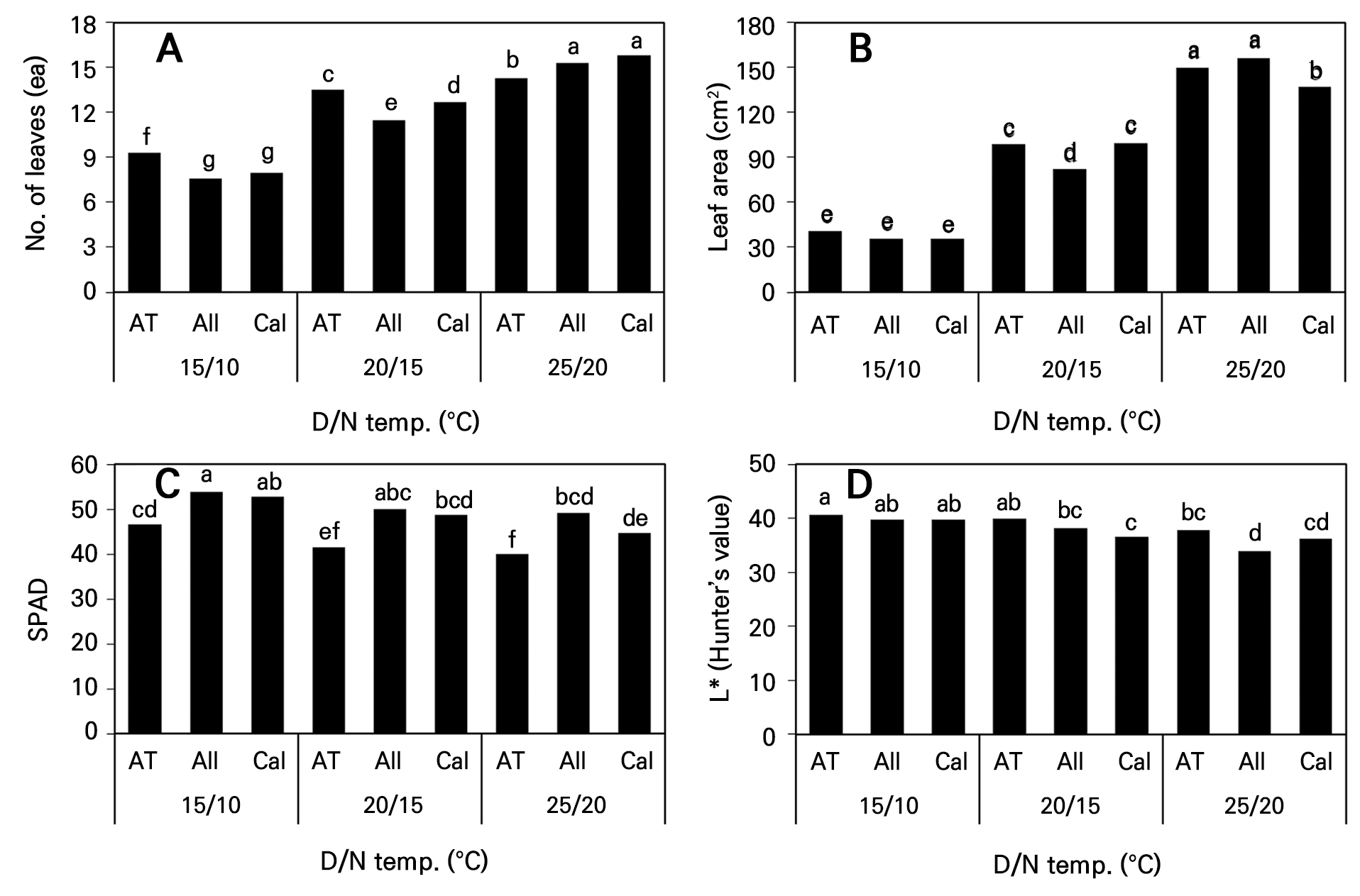
Fig. 4.
Leaf number, leaf area, SPAD, and Hunter L* of three hot pepper cultivars according to day and night temperature changes at 70 days after sowing. Different letters within columns indicate significant difference based on Duncan’s multiple range test, p < 0.05 (n = 3). AT, ‘AT Sinhotan’; All, ‘Allbokhap’; Cal, ‘Callatan’.
본 연구결과와 유사하게 Kang et al.(2005)도 케일 잎의 SPAD 값이 낮을수록 Hunter L* 값이 증가하는 정(+)의 상관 관계를 나타낸다고 본 연구결과와 유사한 보고를 한 바 있다. 실험 종료 시 개화된 식물체는 3 품종 모두 25°C(D)/20°C(N) 처리구에서만 관찰되었으며(Fig. 5) ‘Callatan’ 고추가 1.2개로 많았다(데이터 미제시). 즉 파종 70일 후 조사한 생육조사에서 이미 만개한 상태였으며, 25°C(D)/20°C(N)로 육묘할 경우 육묘기간을 단축시킬 수 있고, 육묘기간 단축을 통해 노동력, 비료 및 관수 비용 등의 생산비 절감 요인이 될 수 있다고 생각한다. Park et al.(2025a)은 양액의 농도를 조절함으로써 고추 육묘기간이 단축될 수 있고, 본 연구자들과 유사한 주장을 한 바 있다.

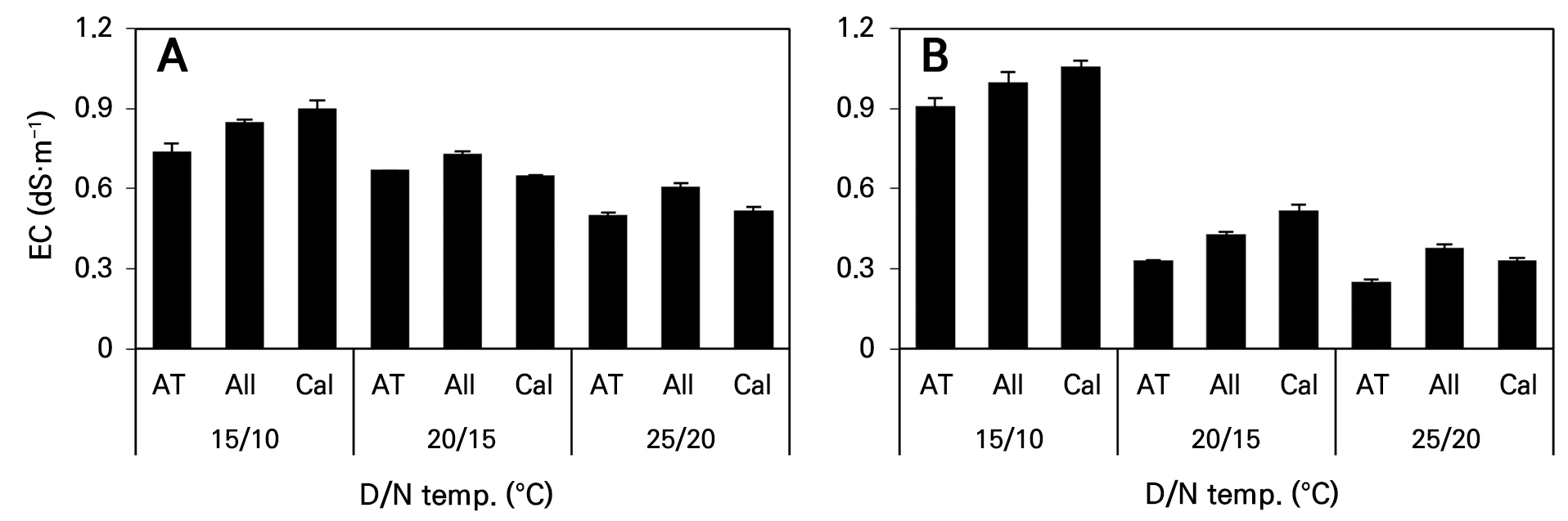
상토의 EC 변화
파종 39일 후 주·야간 온도가 높을수록 세 품종 모두 상토의 EC가 낮아지는 경향이었다(Fig. 6A). 파종 70일 후의 상토 EC도 파종 39일 후의 분석결과와 유사하게 주·야간 온도가 상승할수록 낮아졌으며 모든 품종에서 유사한 경향이었다(Fig. 6B). 특히 온도가 가장 낮게 조절된 15°C(D)/10°C(N) 처리구와 비교할 때 주간온도가 20°C 이상으로 상승하여 식물체의 생육량이 많아질수록 상토의 EC가 50% 이하로 낮아졌다. 이는 15°C(D)/10°C(N) 처리구에서 생장이 가장 저조하여 무기양분 흡수량이 적었지만 주·야간 온도가 상승할수록 생육이 증가하면서 상토에 존재하던 양분의 흡수량이 많아졌고, 파종 39일 후 보다 70일 후의 생장량이 더 많았던 이유로 39일 후의 상토 EC 보다 70일 후의 상토 EC가 낮아졌다고 판단한다. 그러나 15°C(D)/10°C(N) 처리구는 파종 후 39일째 보다 70일 후의 상토 EC가 높았는데, 지상부 생장량이 적은 이유로 인해 온도가 높았던 처리들에 비해 흡수하고 남은 무기원소가 상토에 점차 집적되어 EC가 높은 원인이 되었다고 생각한다. 따라서 생장 온도가 낮게 조절될 경우 양액 공급횟수를 줄이거나 EC를 낮추는 것이 필요하다고 판단한다.
파종 70일 후 잎의 무기성분 함량
파종 70일 후의 엽 조직 내 무기원소 함량을 분석한 결과 주·야간 온도가 높을수록 분석한 대부분의 다량 및 미량원소 함량이 증가하는 경향이었다(Table 1). 높은 온도 처리에서의 생장량 증가와 생장량 증가를 통한 무기원소 흡수량이 증가하여 Fig. 1, Fig. 3 그리고 Fig. 6에 나타낸 바와 같이 상토 EC가 낮아진 원인이 되었다고 생각한다. T-N 함량은 25°C(D)/20°C(N) 처리구의 ‘Allbokhap’과 ‘Callatan’ 고추가 4.0%로 높았으며, P 함량은 20°C(D)/15°C(N) 처리구의 ‘Callatan’ 고추가 3.0%로 가장 높았다. K 함량은 25°C(D)/20°C(N) 처리구에서 높았으며 품종 간 차이는 뚜렷하지 않았다. Yang et al.(2016)은 주·야간 온도차가 토마토 생장에 미치는 영향에 관한 연구결과를 보고한 바 있으며, 주·야간 온도차에 관해 연구한 그들의 연구결과와 주·야간 온도를 모두 상승시킨 본 연구결과를 직접적으로 비교할 수는 없다고 생각한다. 그러나 주간온도가 일정수준까지 높아질 때 지상부 생장량과 무기원소 흡수량이 증가하였다는 그들의 연구결과는 본 연구에서 온도가 상승할 때 생장량과 무기원소 흡수량이 증가한 연구결과에 당위성을 부여한다고 판단한다. 25°C(D)/20°C(N) 처리구에서 각각 ‘Allbokhap’과 ‘Callatan’ 고추가 Ca과 Mg 함량이 높았고, 25°C(D)/20°C(N) 처리구의 ‘Callatan’ 고추가 Cu와 Zn의 함량이 가장 높아 다른 품종보다 흡수율이 높음을 의미한다고 판단한다. Fe 함량은 15°C(D)/10°C(N) 처리구에서 가장 낮았지만 다른 두 온도 처리 그리고 각 온도 처리구 내에서 품종 별 차이가 인정되지 않았다.
