

서언
우리나라의 주요 채소인 배추는 대부분 수확 후 바로 시장에 출하하기 때문에 생산량에 따라 가격의 등락폭이 심하다(Lee and Kang, 1998; Statistics Korea, 2016). 산지의 기후 악화로 인해 배추 생산량의 감소는 배추 가격을 3배 이상으로 급등시켜 배추 소비의 위축으로 직결되었다. 이러한 문제점을 해결하기 위하여 배추의 수급안정화를 위해 배추의 저장이 요구되고 있으며 저장력 향상을 위해 저장 전 처리기술, modified atmosphere package(MAP) 기술 등 다양한 방안들이 제시되고 있다(Bae et al., 2015; Eum et al., 2013). 특히 저장 전 처리기술인 차압예냉은 배추의 수확 후 발생하는 높은 호흡열을 제거시켜 고랭지 여름배추의 저장성을 향상시키는 데 효과적이었다(Eum et al., 2013).
식물체 조직에서 생성되는 에틸렌은 receptor와 결합함으로써 숙성과 노화 등의 다양한 생리활성작용을 조절하는 호르몬이다. 그리고 1-methylcyclopropene(1-MCP)는 에틸렌이 결합해야 할 receptor와 결합함으로써 에틸렌의 활성을 저해시키는 물질로써 에틸렌과는 경쟁관계에 있다(Sisler and Serek, 1997). 그러므로 과일과 채소에서 성숙과 노화를 억제시키는 것이 1-MCP의 주요 기능 중의 하나이다(Cefola et al., 2010; Liu et al., 2013; Meyer and Terry, 2010; Park et al., 2016; Zheng et al., 2014). 1-MCP가 에틸렌의 활성을 억제시키는 농도와 기간은 작물의 종류와 품종 등에 의존한다. 서양배의 경우 품종에 따라 150-500nL·L-1의 다양한 농도로 생리활성을 조절하였다(Li et al., 2013; Wang and Sugar, 2015; Xie et al., 2016). 특히 채소들은 0.1µL·L-1 이하의 낮은 에틸렌 농도에 의해서도 민감하게 반응하는 데 1µL·L-1 1-MCP 처리는 브르콜리의 황변현상과 수확 후 손실을 최소화시키면서 저장기간을 연장시키는 데 효과적이었다(Cefola et al., 2010). Feng et al.(2004)의 연구에 따르면 식물체 조직에서 에틸렌 receptor와 결합한 1-MCP는 영구적으로 결합되어 있으며, 에틸렌의 활성이 회복되는 이유는 조직 내에서 새로운 결합부위가 생성되기 때문이다. 따라서 숙성단계를 거치면서 에틸렌이 계속해서 생성되는 과실과는 달리 배추와 같은 엽채류 작물은 에틸렌 발생량이 적기 때문에 저장 중 1-MCP 처리에 의해서 억제된 에틸렌의 재활성율은 낮다. 그러므로 에틸렌에 의해서 매개된 다양한 생리적 반응들은 억제될 수 있을 것이다.
1-MCP의 또 다른 기능 중의 하나는 호흡률을 저하시켜 품질을 유지시키는 것으로써 사과에서 1-MCP 처리에 의해서 호흡률이 낮아지거나 지연된다고 보고되고 있다(Fan et al., 1999; Fan and Mattheis, 2001). 일반적으로 호흡은 대사활동을 유도하고 증가시킴으로써 저장력을 감소시킨다. 저장 중 호흡의 증가는 원예산물에 특별한 외부의 상처가 없는 상태에서도 시듦과 같은 증상을 유도함으로써 손실률이 증가된다(Watkins, 2006). 배추의 품질을 저하시키는 주요 원인은 수분손실, 경도변화, 그리고 무름병 등이다(Eum et al., 2013). 특히 호냉성 채소인 배추는 6월이후 수확할 경우 높은 외기 온도로 호흡속도는 더욱 빨라지는데, 산지에서 수확 후 상차한 다음 소매지로 유통되는 과정 동안 높은 호흡열에 의해서 품질 저하가 초래된다.
본 연구는 배추의 저온저장을 위한 방안으로 ‘춘광’ 배추에 차압예냉과 함께 1-MCP를 처리함으로써 1-MCP가 저장성에 미치는 영향을 파악하고자 수행하였다.
재료 및 방법
본 연구의 공시재료인 배추 ‘춘광’은 강원도 평창군(해발 550m)에서 재배하여 2016년 6월 14일에 수확한 배추를 이용하였다. 수확된 배추는 즉시 실험실로 옮겨와 초기 품질인자를 조사한 다음 4개의 그룹으로 분류하였다. 4개의 그룹은 차압예냉 처리구, 차압예냉 + Liner 처리구, 차압예냉 + 2µL·L-1 1-MCP 처리구, 차압예냉 + 2µL·L-1 1-MCP + Liner 처리구로 분류하였다. 차압식예냉 처리를 위해서 배추는 농산물 적재 및 유통에 일반적으로 이용되는 플라스틱 박스(52cm×37cm×32cm)에 3-4포기씩 세워서 담아 준비하였다. 배추는 플라스틱 박스를 6개씩(2개×3개) 7단으로 적재한 팔렛트 분량을 본 실험에 이용하였다. 배추의 차압예냉은 5°C에서 차압예냉용 비닐커버를 씌워 간이 차압식 예냉기(FOX-S1004, DSFOX, Korea)를 이용하여 감모율이 3% 이상 되도록 24시간 처리하였다. 수확 직후 배추의 품온은 20°C 내외였고 외기 온도는 25°C 정도였다. 1-MCP 처리는 1-MCP 발생기(FreshLongTM, Ecoplants Co., Korea)를 이용하여 농도가 2µL·L-1이 되도록 설정하여 차압예냉 처리와 동시에 수행하였다. 1-MCP 처리는 자연확산에 의한 처리가 아닌 차압예냉을 진행하는 동안 1-MCP 발생기를 가동시켜 배추 조직으로의 침투를 조장하였다. Liner 처리는 차압예냉 단독 처리 또는 차압예냉과 1-MCP를 동시에 처리한 다음 10cm 간격으로 구멍이 있는 0.03mm linear low density polyethylene(LLDPE, ø 0.03mm) 필름을 이용하여 팔레트 단위로 씌워 저장하였다. 각각의 처리구는 2°C와 상대습도 95% 정도에 저장하였다.
수확 후와 저장 중 실시한 품질평가는 처리구별로 팔레트에서 두 상자씩 선택하여 조사하였다. 저장 중 배추의 무게변화는 저장기간 동안 초기 무게에 대한 감모량을 백분율로 나타내었다. 각종 분석에 이용된 배추 잎은 동일한 조건의 상처가 없는 무결점의 잎을 사용하기 위해 배추 포기를 감싸고 있는 겉잎 1-2엽 바로 아래 잎으로 정하였다. 가용성 고형물 함량은 잎을 착즙한 후 굴절당도계(PAL-1, Atago, Tokyo, Japan)로 측정하여 °Brix로 나타내었다. 경도는 잎의 중륵 부위를 물성분석기(EZ Test / CE-500N, Shimadzu, Kyoto, Japan)를 이용하여 120mm·min-1 crosshead speed 조건으로 직경 5mm probe를 이용하여 측정한 후 N(newton)로 표시하였다.
1-MCP 처리에 따른 수확 후 및 저장시기별 배추의 유전자 발현 정도를 확인하기 위해 SYBR Green 방법을 이용해서 Real time PCR을 수행하였다. Real time PCR을 수행하기 위해 에틸렌 생합성에 관여하는 주요 효소인 1-aminocyclopropane carboxylic acid oxidase(ACO1; AJ309321, ACO2; AJ309322.1), 에틸렌과 결합을 하는 ethylene receptor(ETR1; XM_009106952)와 세포벽 분해효소인 polygalacturonase(PG59; Liang et al., 2015) 유전자들의 primer를 디자인하였다. Total RNA는 배추 잎으로부터 Ribospin Plant kit(GeneAll, Seoul, Korea)를 이용하여 추출한 뒤, Colibri Microvolume Spectrometer(Berthold Titertek Instruments, Inc., Pforzheim, Germany)를 이용하여 각 샘플의 RNA 농도를 확인하였다. 확인된 각 샘플의 RNA로부터 cDNA를 합성하기 위해 Hyperscript RT Premix kit(GeneAll, Seoul, Korea)를 이용하였다. 본 실험에서는 합성된 cDNA를 1/50로 희석해서 PCR template로 사용하였고 Maxima SYBR Green/ROX qPCR Master mix(Thermo Scientific, Waltham, MA, USA)를 이용하여 PCR을 수행하였다. Real time PCR 장비는 PikoReal96(Thermo Scientific, Waltham, MA, USA)을 사용하였다. 유전자 발현 정도는 각 유전자의 standard curve 방법을 사용하며 BrUBC10(Liang et al., 2015) 또는 Ef1α(GO479260, Qi et al., 2010)을 internal reference gene으로 사용하였다.
데이터 분석은 SAS system(SAS Institute Inc. Cary NC 27513, USA)을 이용하여 분산분석(ANOVA)을 하였으며, Duncan의 다중검정(DMRT)을 이용하여 p ≤ 0.05 수준에서 각 시비간의 유의성을 검증하였고, 그래프 작성은 GraphPad Prism5 (Grapgpad Softwaere Inc. La Jolla, CA, USA)을 사용하였다. 모든 처리구는 배추 한 포기씩을 반복구로 하여 항목에 따라 5반복 이상의 실험으로 조사하였다.
결과 및 고찰
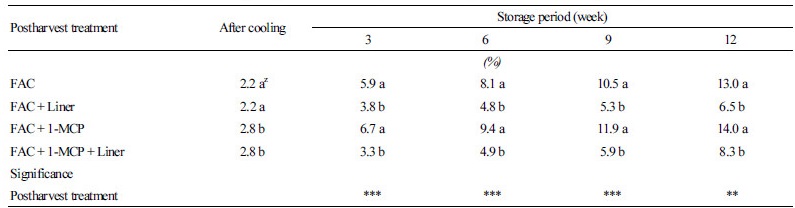
호냉성 채소인 배추의 품질을 저하시키는 주요 원인은 수분손실, 경도변화, 그리고 무름병 등으로 외기 온도가 높을 때 수확이 이루어질 경우에는 배추의 넓은 표면적으로 빠른 호흡 증가와 함께 품질 저하가 초래된다. 이러한 수확 후 호흡증가를 최소화하기 위해서 예냉 기술이 적용되며 이전 연구를 통해서 배추의 차압예냉 처리 효과는 입증되었다(Bae et al., 2015). 배추와 같이 수분을 많이 함유한 원예작물은 배추의 초기무게에 대해서 약 2-3%로 감모율이 진행될 때까지 차압예냉을 실시한다(Eum et al., 2013). 본 연구에서 24시간 동안 차압예냉이 처리된 ‘춘광’ 배추는 약 2.2-2.8%로 감모율이 발생하였다(Table 1). 저장기간 동안 각 실험구의 감모율을 살펴보면 Liner 처리의 유무에 의해서 감모율에 현저한 차이를 보이고 있다. 차압예냉 처리구와 차압예냉 + 1-MCP 처리구는 저장기간 3주가 경과됐을 때 각각 5.9%와 6.7%의 감모율이 발생한 반면 차압예냉 + Liner 처리구, 차압예냉 + 1-MCP + Liner 처리구는 각각 3.8%와 3.3%의 감모율이 발생하였다. 또한 저장기간 12주까지 차압예냉 + Liner 처리구는 감모율이 6.5%였으며, 차압예냉 + 1-MCP + Liner 처리구는 저장기간 9주가 경과되서야 5.9%의 감모율이 발생하였다. 이러한 결과는 배추의 저장을 위해서는 Liner 처리가 반드시 요구되며, 1-MCP 처리는 중량 감모율에 대해서는 영향을 미치지 않는다는 것을 확인할 수 있다. Liner를 처리하지 않으면 저장 9주를 경과하면서 10% 이상의 감모율이 발생되면서 품질이 현저히 저하되었다. 원예작물에서 1-MCP 처리와 저장 중 무게손실률의 상관성은 거의 없다고 보고되고 있으며(Vázquez-Celestino et al., 2016), 오히려 배추와 같은 엽채류의 경우 수확 시 산지에서의 정선손질 과정 동안 발생하는 무게손실은 수확 후 저장 동안 발생하는 손실률에 비해서 더욱 영향을 많이 미친다고 보고되고 있다(Porter et al., 2005).
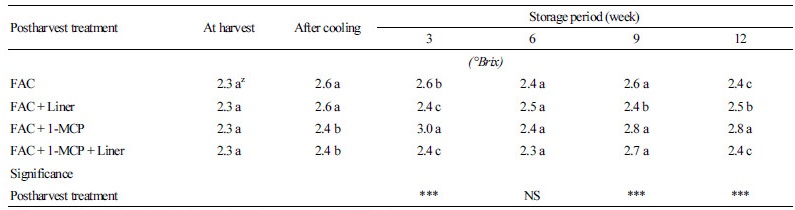
가용성 고형물의 함량은 수확 시에 2.3°Brix였으며, 저장기간 전반에 걸쳐 모든 처리구에서 2.4-3.0°Brix 범위에 포함되었다(Table 2). 각 처리구들은 통계적으로는 유의적 차이를 보이나 값은 0.5°Brix 내외로 저장기간 동안 가용성 고형물의 증감에 따른 경향도 처리 간에 유사하였다. 기존의 보고에서도 1-MCP 처리가 가용성 고형물의 함량에 미치는 영향은 다양하다(Bai et al., 2005). 호흡급등형인 사과에서도 품종마다 차이를 보이며, Gala 품종에서는 가용성 고형물의 손실을 줄인 반면 Delicious, Granny Smith, Fuji 등의 품종에서는 효과가 없었다(Fan and Mattheis, 1999; Fan et al., 1999; Zheng et al., 2014). ‘춘광’ 배추에서 마찬가지로 1-MCP 처리가 가용성 고형물의 함량 변화에 영향을 미치지는 않았다.
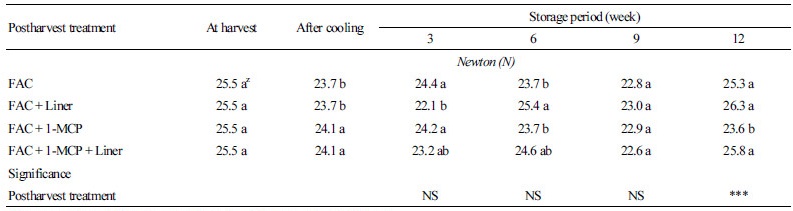
배추의 경도는 수확직후 25.5N이었으며, 차압예냉을 실시한 직후에는 차압예냉과 차압예냉 + Liner 처리구는 23.7N이며 차압예냉 + 1-MCP 처리구와 차압예냉 + 1-MCP + Liner 처리구는 24.1N으로 1-MCP 처리구에서 다소 높게 측정되었다(Table 3). 저장기간 3주가 경과되면서 차압예냉 처리구, 차압예냉 + 1-MCP 처리구는 각각 24.4N, 24.2N으로 Liner 처리구인 차압예냉 + Liner 처리구, 차압예냉 + 1-MCP + Liner 처리구의 22.1N, 23.2N에 비해서 유의적으로 경도가 높았다. 이때의 감모율을 보면 각각 5.9%, 6.7%의 감모율을 보이면서 Liner 처리구에 비해서 약 3% 감모율이 높은 것을 확인할 수 있다(Table 1). 이는 Liner를 처리하지 않아 발생한 감모율이 경도에 영향을 준 것이라 판단된다. 이전 연구결과에 의하면 bell pepper를 이용한 실험에서도 4주간의 저장기간 동안 필름 무처리구는 필름 처리구에 비해서 5% 이상의 수분손실률을 야기시켰는데, 이는 직접적인 경도의 증가로 이어졌다(Ben-Yehoshua and Rodov, 2003). 또한 Liner 처리구들에서도 1-MCP 처리구는 경도가 유의적으로 높았다. 그러나 저장기간 6주가 경과되면서 처리들 간에 경도 변화는 무게 감소율과 뚜렷한 경향을 보이지 않았으며, 저장기간 9주까지 서서히 감소하였다. 저장기간 12주가 경과되면서 다시 증가하는 경향을 보이는데 감모율이 13-14%까지 발생하였음에도 불구하고 경도는 배추를 수확직후 측정한 경도인 25.5N과 비교해 볼 때 차압예냉, 차압예냉 + Liner, 그리고 차압예냉 + 1-MCP + Liner 처리구에서 각각 25.3N, 26.3N, 그리고 25.8N으로 초기의 경도와 큰 차이를 보이지 않고 있다. 이러한 현상은 배추를 저장할 경우 저장일수가 경과되면 배추의 제일 바깥에 있는 배춧잎은 제거를 하게 된다. 이러한 경우 발생하는 손실률을 정선손실률이라 하는데 기존 결과에서 ‘춘광’ 배추의 정선손실률은 저장 6주가 경과 되면서 약 12%의 손실률이 발생되었다(Eum et al., 2013). 이처럼 정선에 의해서 배추의 겉잎을 제거한 다음에 경도를 측정하면 결구가 형성되는 배추와 같은 엽채류의 경우 처리 간 경도의 유의적 상관관계를 확인하는 것은 어렵다고 판단된다. 결국 1-MCP 처리는 배추를 저장하는 하는 동안 ‘춘광’ 배추의 외관상 품질에는 영향을 미치지 않았으므로 1-MCP가 배추의 숙성에 관여하는 유전자 발현과의 상관성에 대해서 확인하였다.
1-MCP의 활성은 원예작물의 품목, 품종 그리고 조직에 영향을 받는다. 어떤 원예작물의 1-MCP의 효과를 얻는 데 높은 농도가 요구된다면 이는 성숙 또는 숙성이 일어나는 조직에서 새로운 receptor가 계속해서 생성되기 때문에 혹은 조직에 존재하는 receptor와 처리된 1-MCP의 친화력이 약하기 때문에 1-MCP의 농도 혹은 처리기간이 길어질 수 있다고 판단된다(Sisler and Serek, 1997). 1-MCP가 receptor와 잘 결합하기 위해서는 온도가 주요 요인이다. 바나나의 경우 1-MCP 처리온도는 2.5°C보다 15-20°C에서 효과적이라고 보고된다(Jiang et al., 2002). 엽채류인 양상추의 경우도 20°C에서 저장할 품질이 급격히 저하되지만 1-MCP를 처리하면 갈변 억제, 가용성 당 및 단백질 함량 유지 등의 효과가 있다(Tian et al., 2014). 저장을 목적으로 수확하는 원예작물은 일반적으로 바로 출하되는 작물에 비해서 약 80% 정도의 성숙이 진행된 시점에서 수확이 이루어지는데 특히 품질 저하가 빠르게 진행되는 엽채류를 유통할 경우 농가에서는 배추의 크기가 판매 가능한 크기에 도달하면 수확하며, 저장을 실시할 경우에도 이와 유사하다(Bae et al., 2015). 특히 배추는 저장 전 처리기술인 차압예냉 처리에 의해서 저장기간을 연장할 수 있다. 일반적으로 배추의 차압예냉은 2°C에서 처리되지만, 기존 1-MCP 처리시 2°C에서 효과가 없었으므로(자료 미제시) 본 연구는 1-MCP의 효과를 확인하기 위해 5°C에서 이루어졌다. 기존 실험에서 배추에서의 차압예냉 효과는 0-8°C 범위 내에서 유사하였다(자료 미제시). ‘춘광’ 배추에서 1-MCP의 효과 유무를 관찰하기 위해서는 더 온도를 높여야 하겠으나, 저장을 목적으로 한 배추의 경우 5°C보다 높은 온도에서의 처리는 오히려 배추의 저장 전 초기 품질저하를 초래하여 저장기간 및 저장력을 저하시킬 수 있다고 판단된다.
에틸렌 생합성에 관여하는 주요 효소는 1-aminocyclopropane carboxylic acid oxidase(ACO)와 1-aminocyclopropane carboxylic acid synthase(ACS)이며 에틸렌은 ethylene receptor(ETR)와 결합함으로써 생리활성을 조절한다. 이와 관련하여 1-MCP처리에 대한 ACO 및 ETR 유전자들의 발현 정도를 확인하였다. 배추에 존재하는 ACO 유전자는 두 종류로 BrACO1과 BrACO2 유전자이다(Fig. 1). 각 처리구 그룹별(차압예냉 처리구, 차압예냉 + Liner 처리구, 차압예냉 + 2µL·L-1 1-MCP 처리구, 차압예냉 + 2µL·L-1 1-MCP + Liner 처리구)간 BrACO1과 BrACO2 유전자 발현양상을 비슷하였다. 차압예냉을 실시한 직후에는 유전자의 발현량이 많다가 저장 3주가 경과하면서 감소된 유전자 발현은 저장 기간이 경과되면서 증가하였다. 이러한 결과는 ETR 유전자의 발현에도 유사한 경향을 보였다(Fig. 2). 배추에 존재하는 BrETR1 유전자는 ACO 유전자들과는 달리 차압예냉 처리 직후에는 유전자의 발현량이 적다가 저장기간이 경과되면서 증가하였다. 그러나 1-MCP 처리유무는 저장기간 동안 처리구 그룹별 BrETR1 유전의 발현에는 통계적으로 유의한 영향을 미치지 않았다. 결과적으로 ‘춘광’ 배추에서는 1-MCP 처리가 에틸렌 생합성과 관련된 유전자인 ACO와 ETR 유전자 발현 양상에는 영향을 미치지 않는다는 것을 의미한다. 기존 보고에서도 배추에 0.1µL·L-1와 1.0µL·L-1로 처리농도를 달리하여 1-MCP를 처리할 경우 처리 직후에는 대조구에 비해서 호흡률이 감소하지만 저장 1주일이 경과하면서 대조구와 차이를 보이지 않았으며 이러한 경향은 저장 9주까지 지속되었다(Porter et al., 2005). 에틸렌 발생률에서 유사한 경향을 보이는데, 1-MCP를 처리한 직후에는 호흡률과 반대로 대조구에 비해서 급격한 에틸렌의 증가를 보이다가 저장1주일이 경과된 시점에는 대조구와 유의적 차이를 보이지 않았다. 이러한 현상은 본 실험결과와 유사성을 보이는데 2.0µL·L-1 1-MCP를 처리한 직후 에틸렌 생합성 효소인 ACO 유전자의 일시적인 증가가 나타난 후 저장기간이 경과되면서 감소하면서 1-MCP 처리유무에 의한 차이는 없었다(Fig. 1).
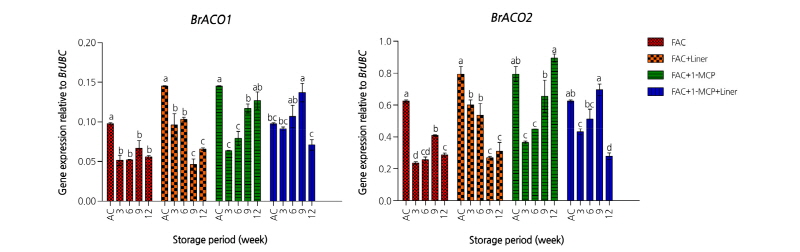
Fig. 1. Expression levels of 1-aminocyclopropane carboxylic acid oxidase (BrACO1, BrACO2) genes on spring Kimchi cabbages ‘Choongwang’ treated with force air cooling (FAC) and affected by different condition (with/without 2 µL·L-1 1-MCP, with/without Liner of 0.03 mm LLDPE perforated film) during storage at 2°C for 12 weeks. AC represents after cooling. All the values are expressed as means ± SE of three replicates. The different letters indicate significant differences during storage period at the 5% level for Duncan’s multiple range test.
결과적으로 비호흡급등형인 배추에서는 1-MCP 처리에 의한 에틸렌 생합성을 줄이는 데 효과가 미비하다고 판단된다. 복숭아 등의 과실에서 1-MCP 처리는 에틸렌 생합성과 관련된 유전자의 발현을 조절함으로써 에틸렌 생성량은 억제하며, 아보카도의 경우에는 pre-climacteric 단계에 1-MCP를 처리함으로써 ACS와 ACO의 활성과 유전자 발현을 억제한다고 보고된 바 있다(Bregoli et al., 2005; Defilippi et al., 2002; Owino et al., 2002). 비호흡급등형 고추에서도 색발현이 시작되는 숙성단계에 처리된 1-MCP는 숙성을 지연시키고, 에틸렌 생합성에 관여하는 유전자(CaACO3, CaACO5, CaACS1, CaETR4, CaETR5)의 발현을 억제시켰다(Aizat et al., 2013). 그러나 녹과 상태에서는 그 효과가 미비하거나 없었다.
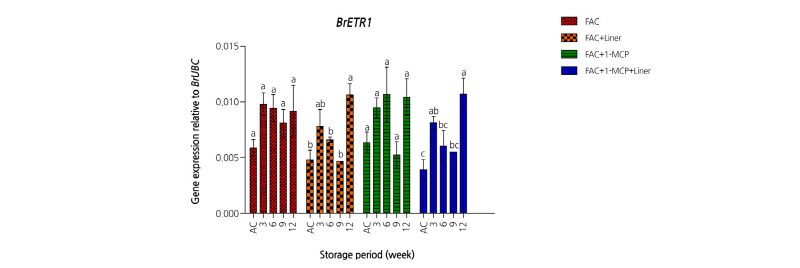
Fig. 2. Expression levels of ethylene receptor (BrETR1) gene on spring Kimchi cabbages ‘Choongwang’ treated with force air cooling (FAC) and affected by different condition (with/without 2 µL·L-1 1-MCP, with/without Liner of 0.03 mm LLDPE perforated film) during storage at 2°C for 12 weeks. AC represents after cooling. All the values are expressed as means ± SE of three replicates. The different letters indicate significant differences during storage period at the 5% level for Duncan’s multiple range test.
1-MCP의 다른 주요 기능 중의 하나는 세포벽 대사에 관여함으로써 원예작물의 성숙 및 노화 과정 동안 발생하는 연화를 지연시킨다. 1-MCP가 처리된 바나나에서 연화가 지연되었는데 이는 세포벽 분해효소인 pectin methylesterase와 polygalacturonase (PG), endo-b-1,4-glucanase 등의 활성이 1-MCP에 의해서 낮아졌기 때문이다(lohani et al., 2004). 반면 딸기나 오렌지와 같은 비호흡급등형 과실에서 1-MCP 처리는 연화를 억제하는 데 효과가 없었다(Porat et al., 1999; Tian et al., 2000). PG는 배추 세포벽의 주성분인 pectin을 분해하며, 이러한 현상에 의해서 세포벽의 구조적인 변화가 초래되어 잎 조직의 연화가 발생할 수 있다. 이와 관련하여 배추에서 1-MCP 처리 유무에 대한 PG 유전자의 발현 정도를 확인하였다. 배추에서 존재하는 PG 유전자는 BrPG59이며, 차압예냉 후 처리그룹별 PG 유전자의 활성은 처리 간에 다소 차이를 보인다. Liner 무처리구는 저장기간 전반에 걸쳐 PG 유전자의 발현이 저조한 데 비해, Liner 처리구에서는 저장기간이 경과함에 따라 PG 유전자의 발현이 증가하는 양상을 보이며 BrPG59 유전자의 상대적 비율이 높았다(Fig. 3).
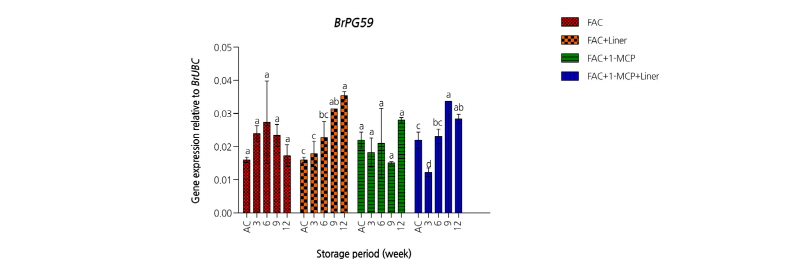
Fig. 3. Expression levels of ethylene polygalacturonase (BrPG59) gene on spring Kimchi cabbages ‘Choongwang’ treated with force air cooling (FAC) and affected by different condition (with/without 2 µL·L-1 1-MCP, with/without Liner of 0.03 mm LLDPE perforated film) during storage at 2°C for 12 weeks. AC represents after cooling. All the values are expressed as means ± SE of three replicates. The different letters indicate significant differences during storage period at the 5% level for Duncan’s multiple range test.
이상의 결과를 종합해 보면 무게변화, 당도, 경도 등의 품질 인자와 PG, ACO, ETR 의 유전자 발현양상을 기준으로 배추를 평가할 경우, 1-MCP 처리는 2°C의 저온저장이 요구되는 배추에서는 효과가 미비하다고 판단된다. 하지만 저장기간 동안 ‘춘광’ 배추의 무게손실을 최소화 하기 위해서는 PE film을 이용한 Liner 처리가 반드시 요구되며 이를 통해 처리 후 9주까지 품질유지가 가능하였다.
