

서 언
산업혁명 이후 화석연료의 사용이 급증하면서 연소 시 발생하는 이산화탄소 등 온실가스의 대기 중 농도가 급격히 증가하고 있으며, 이로 인해 지구 밖으로 방출되는 복사열이 감소하여 대기온도가 상승하는 지구 온난화 현상이 전 세계적으로 문제되고 있다. 기후변화에 관한 정부간 패널(Intergovernmental Panal on Climate Change, IPCC)의 4차 보고서에 따르면 산업혁명 이전에 280µL・L-1이었던 대기 중 이산화탄소 농도가 현재는 381µL・L-1로 증가되었으며, 이로 인해 전 세계적으로 대기 온도가 지난 100년간 0.74°C 상승하였다고 보고되었다(IPCC, 2007). 우리나라는 근대적인 기상관측이 시작된 1904년부터 2000년까지 약 100년 동안 평균기온이 약 1.5°C 상승하여 전 지구적인 온난화 추세를 상회하고 있으며(Kwon et al., 2007), IPCC의 온실가스 배출 시나리오 중 A1B 기후변화 시나리오를 적용했을 때 21세기 후반에 대기 중 이산화탄소 농도는 약 700µL・L-1로 상승하며, 대기온도는 약 1.8-4.0°C 정도 증가할 것으로 예측되고 있다(Korea Meteorological Administration, 2009).
온도는 식물의 생리・생태에 커다란 영향을 미치며, 특히 원예작물의 경우 생육기 온도에 의해 생산량과 품질이 크게 좌우되기 때문에 중요한 환경요인으로 인식되고 있다(Florides and Christodoulides, 2009; Lee et al., 2008). Zamski and Schaffer(1996), Lakso and Seeley(1978)는 생육 적온을 초과하는 온도조건 하에서 원예작물은 광합성률이 저하하는 반면, 호흡률은 증가하기 때문에 순광합성률이 감소하여 수체 생육 및 품질이 크게 저하된다고 하였다(Faust, 1989; Tomana and Yamada, 1988; Saure, 1990). 반면에 대기 중 이산화탄소 농도가 증가하면 이산화탄소 분압이 높아져 대기에서 식물체 내 광합성 기관까지 이산화탄소가 쉽게 확산될 뿐 아니라, 광합성 시 이산화탄소 고정효소인 rubisco의 반응이 촉진된다(Kimball et al., 2002). 따라서 이산화탄소에 민감하게 반응하는 C3 식물을 대상으로 한 일련의 연구에서 대기 중 이산화탄소 농도를 2배 정도 증가시킬 경우 광합성이 약 25-75% 정도 증가한다고 보고되어 있다(Luxmoore et al., 1993; Urban, 2003).
포도는 국내 과수산업의 약 20%를 차지하는 대표적인 작물로 온도적응성이 비교적 넓은 작물로 알려져 있으나(Buttrose, 1969), 품종에 따라 온도에 민감하게 반응하여 수체생육 및 과실품질에 심각한 피해를 받을 수 있다(Lee et al., 1979). 그런 이유로 온도 등 환경요인에 대해 많은 연구가 수행되었으나, 기후변화 시나리오를 기반으로 온도와 이산화탄소의 조합처리가 포도의 수체생육 및 과실품질에 미치는 영향을 구명한 연구는 미흡한 실정이다.
따라서 본 연구는 A1B 기후변화 시나리오를 적용한 온도(+ 4.0°C)와 대기 중 이산화탄소 농도(700µL・L-1)를 국내 포도 주품종인 ‘캠벨얼리’에 조합 처리함으로써 수체생장 및 과실품질 변화에 미치는 영향을 구명하는 한편, 기후변화에 대응할 수 있는 재배법 개발에 필요한 기초 자료를 확보하기 위해 수행하였다.
재료 및 방법
실험재료 및 처리
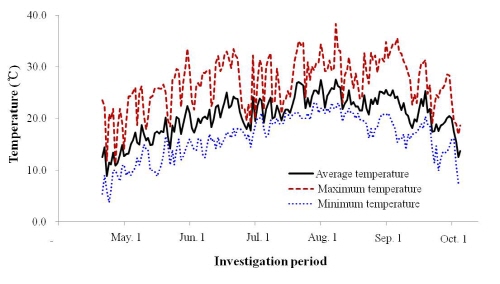
경기도 수원시 이목동에 위치한 국립원예특작과학원 내 온도 및 이산화탄소 제어가 가능한 독립된 4개의 라이조트론에 타발휴면 중인 수세가 일정한 3년생 ‘캠벨얼리’ 포도를 처리구당 5주씩 정식하여 실험을 수행하였다. 처리구는 A1B 시나리오를 바탕으로 온도 및 이산화탄소 농도에 따라 대조구(대기온도, 390µL・L-1CO2), 이산화탄소 상승구(대기온도, 700µL・L-1CO2), 온도 상승구(대기온도 + 4.0°C, 390 µL・L-1CO2), 이산화탄소 + 온도 상승구(대기온도 + 4.0°C, 700µL・L-1CO2)로 각각 설정하였다. 라이조트론 내 환경조절은 실시간으로 수집된 기상정보를 바탕으로 중앙분산식 제어시스템(POREX 6800, POSCON Co., Korea)으로 온도를 조절하였고, Infra-red 가스 분석기(ZRH model, Fuji Electric Co., Japan)를 이용하여 이산화탄소 농도를 조절하였다. 실험기간은 2011년 4월 21일부터 9월 1일까지 총 134일간에 걸쳐 수행하였으며 실험기간 중 기온변화는 Fig. 1과 같다.
수체 생육 및 과실품질 조사
신초 생육은 대조구의 수확이 종료된 2011년 9월 1일에 신초길이, 직경 및 부초수를 조사하였다. 과실은 착색도를 기준으로 처리구별로 수확하여 과립중 및 종・횡경을 측정하였으며, 처리구당 20립의 과립을 무작위로 선정하여 물성분석기(GB/TAXT Express 440, Surrey, UK)로 과립 경도를 측정하였다. 당함량은 과실의 과즙을 취해 디지털 당도계(PR-101, Atago, Japan)로 측정하였으며, 산함량은 여과한 과즙 5mL에 증류수 20mL를 가한 후 0.1N NaOH로 적정하면서 pH 7.0이 될 때까지 소요된 NaOH량을 tartaric acid량으로 환산하였다. 안토시아닌 함량은 과육을 제거한 과피를 1g 채취해 10% Fomic acid/MeOH에 24시간 추출한 후, 추출액 1mL에 0.2M sodium acetate(pH 1.0) 9mL를 가한 혼합액을 HPLC(1525, Waters, USA)를 이용해 520nm로 측정하였다. 안토시아닌 정량을 위해 malvidin-3-glycoside를 표준물질로 사용하였다. 과피의 착색도 변화는 포도 칼라챠트를 이용해 변색이 시작된 7월 26일부터 9월 1일까지 총 12회 조사하였다.
광합성 및 증산량 측정
광합성률과 증산량의 경시적 측정은 전엽 30일 후의 건전한 성엽을 대상으로 광합성 측정기(LI-6400, Li-Cor, USA)로 6월 20일부터 9월 15일까지 2주 간격으로 측정하였다. 측정 조건은 PPDF(photosynthetic photon flux density)를 1000μmol・m-2・s-1 로 설정하였으며, 측정 시 챔버의 온도와 이산화탄소 농도는 각각의 처리조건과 동일하게 설정하여 측정하였다.
결과 및 고찰
수체생육 및 생리반응
기후변화 시나리오에 따른 온도 및 이산화탄소 처리가 ‘캠벨얼리’ 포도의 수체생장에 미치는 영향은 Table 1과 같다. 생육기 중 신초 길이를 조사한 결과, 대조구와 이산화탄소 상승구는 각각 206.2, 224.8cm로 통계적으로 유의차 없이 처리구 중 가장 낮은 수치를 보인 반면, 온도 상승구(255.6cm), 이산화탄소 + 온도 상승구(312.6cm) 순으로 신초 길이가 증가하여 온도의 증가에 의해 신초 신장이 왕성해진 것을 알 수 있었다. 생육기 중 부초 발생량은 대조구와 이산화탄소 상승구에서는 3.4, 3.0개가 발생한 반면, 온도 상승구와 이산화탄소 + 온도 상승구에서는 6.2, 5.2개가 발생하여 온도와 신초의 영양생장 간에 고도의 상관관계가 인정되었다. 하지만 신초 직경은 온도 상승구와 이산화탄소 + 온도 상승구가 각각 5.96, 6.35mm로 대조구와 이산화탄소 상승구의 7.41, 7.56mm보다 감소한 것으로 나타나 신초 생장량 및 부초 발생량과 상반되는 결과를 보였다. Buttrose(1969)는 온도에 따른 포도의 신초장 변화를 조사한 결과, 25-30°C의 온도 범위에서 가장 높았다고 보고하였는데 본 실험에서는 발아 후 60일 사이인 6월부터 7월 중순까지의 일 평균온도 범위가 17.6-27.2°C이었기 때문에 이보다 4°C 높게 설정한 온도 상승구와 이산화탄소 + 온도 상승구가 양・수분 흡수 및 대사작용이 활발하여 신초 신장량이 증가한 것으로 생각된다(Kirschbaum, 2004).
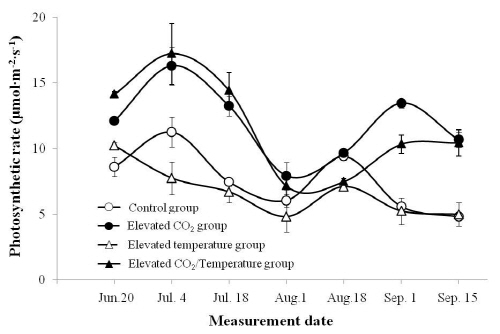
생육기 중 광합성률의 경시적 변화를 조사한 결과(Fig. 2), 이산화탄소 상승구와 이산화탄소 + 온도 상승구의 광합성률은 생육기 동안 높은 광합성률을 보였다. 이는 영년생 작물인 포도와 목화에 고농도의 이산화탄소 처리를 했을 때 광합성률과 생산량이 증가한다는 Kimball et al.(2002)의 실험 결과와 동일하였다. 본 실험에서도 식물체의 광합성과 생체중 생산에 필수적 역할을 하는 대기 중 이산화탄소의 농도가 증가하면서 포도 잎의 광합성 기관까지 이산화탄소의 확산 및 이용이 용이해졌기 때문에 광합성률이 증대되었다고 생각되었다(Florides and Christodoulides, 2009; Kimball et al., 2002; Urban, 2003). 하지만 7월 중순부터 8월 중순까지 이산화탄소 상승구와 이산화탄소 + 온도 상승구가 대조구에 비해 광합성률이 급격히 감소하는 것으로 보았을 때 고온조건에 의해 이산화탄소의 광합성 증대효과가 억제된 것이라고 추측되었다. 특히 이산화탄소 + 온도 상승구의 경우 광합성률이 현저히 감소하여 8월 18일에는 대조구보다 낮은 광합성률을 나타냈는데 이는 8월 중 온도가 35°C에 육박했기 때문에 대기온도보다 4°C 높게 설정한 이산화탄소 + 온도 상승구는 고온에 의해 이산화탄소의 고정속도 및 광계 II계의 활성이 저해되었을 뿐 아니라(Havaux et al., 1991; Mooney et al., 1978), 엽록소 등 광합성과 관련한 기관이 장해를 받았기 때문이라 생각되었다(Zamski and Schaffer, 1996).
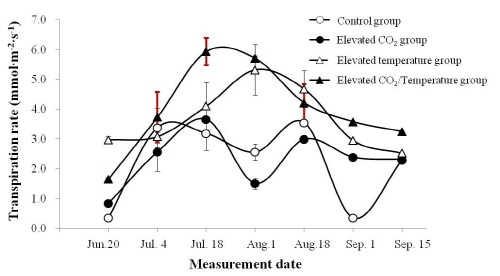
생육초기의 증산량은 처리구간에 큰 차이를 보이지 않았으나(Fig. 3), 하계의 고온이 지속되면서 이후 생육기간 동안 온도 상승구와 이산화탄소 + 온도 상승구의 증산량이 대조구와 이산화탄소 상승구보다 높았다. 이는 온도 상승구와 이산화탄소 + 온도 상승구가 대조구와 이산화탄소 상승구보다 4°C 높게 설정된 고온 조건이기 때문에 잎과 대기 사이에 높은 증기압차가 발생하여 증산량이 상승한 것으로 생각된다(Kirschbaum, 2000, 2004). 다만 이산화탄소 농도에 따른 증산량의 변화는 일정한 경향을 보이지 않아 고농도의 이산화탄소처리 시 기공전도도의 감소 및 기공폐쇄에 의해 증산량이 감소한다는 일련의 연구결과와 차이가 있었다(Allen, 1990; Medlyn et al., 2001). 또한 Kirschbaum(2004)에 의하면 기후변화 시 일반적으로 태양광량 및 대기 중 상대습도가 증가되는 등 기타 환경요인이 추가적으로 작용하기 때문에 증산량의 정확한 구명을 위해서는 여러 환경요인을 대상으로 구체적 연구가 필요하다고 생각되었다.
과실품질 및 생리반응
상승온도와 이산화탄소가 ‘캠벨얼리’ 품종의 과실특성 및 생육단계에 미치는 영향은 Table 2와 같다. 과립중 및 과립의 종경은 온도 및 이산화탄소 농도 간 유의성이 인정되지 않았으나, 횡경은 대조구가 15.6mm로 가장 작은 수치를 보였으며, 온도 상승구(17.4mm), 이산화탄소 상승구(17.7mm), 이산화탄소 + 온도 상승구(18.1mm) 순으로 증가하여 온도 및 이산화탄소의 농도가 높을수록 증가하는 경향을 보였다. 온도 상승구에서 횡경이 증가한 것은 포도의 생육기 중 주야간 온도가 25-35°C 범위에서 고온일수록 과실비대가 우수하였다는 Kliewer(1977)의 연구와 유사한 결과로 한계온도를 초과하지 않는 범위 내에서 고온일수록 세포비대가 왕성해져 세포크기가 증가하였기 때문이라고 생각되었다(Adams et al., 2001; Sugiura, 1997). 특히 이산화탄소 + 온도 상승구의 경우 과립비대 초기의 고온에 의해 세포비대가 우수했을 뿐 아니라, 과실비대기 동안 고농도의 이산화탄소에 의해 광합성률이 상승하여 다량의 동화산물을 생산하여 과립에 전류되었기 때문에 과립비대에 유리하였을 것으로 판단된다. 각 처리구의 생물계절 변화는 대조구의 만개일은 6월 11일, 수확일은 9월 1일이었던 것과 비교해 이산화탄소 상승구에서는 만개일과 수확일이 각각 5일과 4일, 온도 상승구에서는 7일과 2일, 이산화탄소 + 온도 상승구에서는 10일과 11일이 단축되는 등 지구온난화에 의해 ‘캠벨얼리’ 포도의 수확기까지의 생물계절이 단축될 것으로 전망되었다.
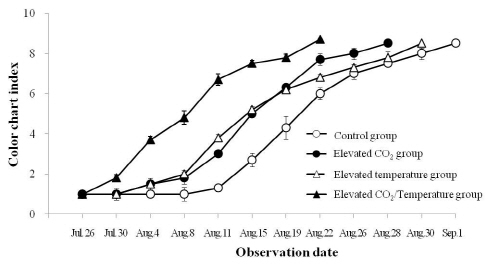
‘캠벨얼리’ 포도의 당함량은 이산화탄소 상승구가 14.6 °Brix로 처리구 중 가장 높은 수치를 보인 반면, 온도 상승구가 13.9°Brix로 가장 낮은 수치를 보여 당함량과 대기 중 이산화탄소 농도와 높은 상관관계가 있음을 알 수 있었다(Table 3). 이는 토마토를 이용한 실험에서 고농도의 이산화탄소를 처리했을 때 효소활성 및 당함량이 증가한다는 Islam et al.(1996)의 결과와 일치하였다. 과피의 안토시아닌 함량은 처리구간 유의성이 인정되지 않았는데 이는 처리구별 수확시기를 달리했기 때문이라 생각된다. 하지만 칼라챠트를 이용해 포도의 착색도를 조사한 결과(Fig. 4), 착색개시일이 이산화탄소 + 온도는 7월 30일, 온도 상승구와 이산화탄소 상승구가 8월 4일, 대조구가 8월 11일로 이산화탄소 + 온도 상승구가 대조구보다 약 12일 빠르게 착색되어 고온에 의해 착색이 지연된다는 기존의 연구결과(Kliewer, 1977; Lee et al., 1979)와 상반되는 결과를 얻었다. Yamane et al.(2006)는 포도의 변색초기에 고온이 안토시아닌의 합성 및 축적을 크게 저하한다고 보고하였는데 본 연구에서는 이산화탄소 + 온도 상승구가 생육기간 동안 높은 광합성률에 의해 동화산물의 축적이 많았을 뿐 아니라, 생물계절이 앞당겨졌기 때문에 연중 최고기온을 보였던 8월 중순에는 착색이 상당수 진행되어 고온에 의한 착색저하 현상을 회피하였다고 생각되었다.
|
Fig. 4. Changes of pricarp colouration according to different conditions of CO2 and temperature in ‘Campbell Early’ grapevines. Bars represent the standard error of means from 10 replications. |
이와 같은 결과를 종합하면 미래 기후변화 시 ‘캠벨얼리’ 포도는 신초 길이 및 부초수 증가 등 생장은 크게 증가할 것으로 생각되나, 광합성률의 감소로 신초직경 등 수체의 생육 충실도는 전반적으로 저하될 것으로 예측된다. 반면에 대기 중 이산화탄소 농도의 증가로 과실 당함량이 증가할 뿐 아니라 온도상승에 의해 생물계절이 단축되면서 변색기가 앞당겨져 8월 중순의 최고기온을 회피하기 때문에 착색장해를 크게 받지 않을 것으로 예측되었다. 하지만 영년생인 포도의 경우 앞에서 언급한 불충실한 수체 생육이 누적되면서 과립의 당함량 및 착색이 저하될 가능성이 있으므로 상승온도 및 이산화탄소의 장기처리에 관한 연구가 추가적으로 수행될 필요가 있다고 생각되었다.
