

Introduction
Materials and Methods
Plant material and virus diagnosis
In vitro culture establishment and experimental design
Thermotherapy and ribavirin treatments
Shoot-tip culture and plant regeneration
Data collection and statistical analysis
Results
Effects of thermotherapy and ribavirin treatments on the in vitro shoot growth of grapevines
Changes in shoot-tip regeneration and plant survival under single and combined treatments
Comparison of GLRaV-3 and GFkV elimination efficiency rates under different treatment conditions
Discussion
Conclusion
Introduction
Viral diseases that affect grapevines (Vitis vinifera L.) are significant constraints in viticulture worldwide, leading to reduced vegetative growth, yield losses, and a decline in fruit quality. These challenges threaten the long-term sustainability of grape and wine production systems (Krenz et al. 2023; Cieniewicz and Fuchs 2025). As plant viruses are easily spread through vegetative propagation and insect vectors, producing and distributing virus-free propagation materials is crucial for maintaining competitiveness in grapevine cultivation systems that rely heavily on clonal propagation (Gao and Lozano-Durán 2025). Among grapevine viruses, Grapevine leafroll-associated virus 3 (GLRaV-3) is the most widely distributed and is considered a major pathogen, responsible for reduced vine vigor, delayed fruit maturation, yield decline, altered berry composition, and compromised wine quality (Hančević et al. 2022; Fust et al. 2025). Additionally, Grapevine fleck virus (GFkV) has been consistently reported in older vineyards and rootstock–scion germplasm collections, where infection is generally associated with chlorotic flecking along the leaf veins, mild interveinal yellowing, reduced vegetative growth, and weakened vine vigor (Basso et al. 2017; Thapa et al. 2024). Recent surveys in Korea have confirmed a high incidence of co-infection by GLRaV-3 and GFkV in several commercial cultivars (Kim et al. 2021; Choi et al. 2024). Notably, the table grape cultivar ‘Shine Muscat’, which holds significant economic importance, shows a high frequency of mixed infection of these two viruses, underscoring the growing demand for reliable virus-free planting materials to ensure stable production (Yu et al. 2023; Ren et al. 2024).
Co-infections of GLRaV-3 and GFkV reportedly exert more severe negative effects on vine growth and productivity than single-virus infections (Nuzzo et al. 2022; Salo et al. 2024). When both viruses are present simultaneously, the characteristic symptoms associated with each pathogen—such as leaf discoloration, leaf rolling, and chlorotic flecking along the veins—may overlap, potentially resulting in a greater reduction of the photosynthetic capacity and disruption of assimilate partitioning. These physiological disturbances may further exacerbate shoot growth suppression and vine weakening, leading to delayed fruit maturation, reduced berry size, uneven coloration, and instability in the sugar–acid balance of the fruit. Several studies have suggested that co-infection also promotes increased viral accumulation or earlier symptom expression, thereby posing a long-term threat to vineyard productivity and stability (Sadras et al. 2024). Thus, GLRaV-3 and GFkV co-infection may intensify disease severity not only through their simultaneous presence but also through additive or synergistic interactions.
Given the severity of such mixed infections, the development of highly efficient sanitation techniques has become a critical issue in the grape industry. Currently, shoot-tip culturing combined with thermotherapy is the most widely adopted sanitation method for grapevines (Wang et al. 2018; Hu et al. 2020). More recently, strategies incorporating ribavirin-based chemotherapy or shoot-tip cryotherapy have been explored to improve the efficiency of virus elimination (Zhang et al. 2024). However, grapevine responses to sanitation treatments vary considerably among cultivars, and success with regard to plant survival and virus elimination depends on multiple factors, including the intensity and duration of the thermotherapy, the composition of the culture medium, and the concentration of antiviral agents (Wante et al. 2026). In particular, under co-infection conditions, the differential biological responses of individual viruses may limit the effectiveness of sanitation systems originally designed for single-virus infections. Therefore, the establishment of optimized sanitation protocols specifically tailored to GLRaV-3 and GFkV co-infection is necessary.
This study applies ribavirin (20 and 40 mg L-1), thermotherapy at 37°C, and treatments that combine these two strategies alongside shoot-tip culturing to V. vinifera ‘Shine Muscat’ plants co-infected with GLRaV-3 and GFkV. By comparing the effects of each treatment on in vitro growth, regeneration performance, and mixed virus elimination efficiency, this research aims to establish an optimized sanitation protocol for the effective production of virus-free ‘Shine Muscat’ planting materials.
Materials and Methods
Plant material and virus diagnosis
This study was conducted in April of 2022 during the active shoot-emergence stage using three-year-old Vitis vinifera L. ‘Shine Muscat’ vines grown in a private commercial vineyard located in Gimhae, Gyeongsangnam-do, Republic of Korea (36.1398° N, 128.1136° E). For the experiment, young elongating shoots showing clear virus-like symptoms, including leaf rolling, chlorosis or flecking along the veins, foliar discoloration, and an overall reduction in vine vigor, were selected. These symptom criteria were based on characteristics commonly reported in relation to GLRaV-3- and GFkV-infected grapevines (Basso et al. 2017; Hančević et al. 2022; Thapa et al. 2024). Actively growing shoots exhibiting clear viral symptoms were collected for the experiment. The vineyard is situated in the warm temperate climatic zone of southern Korea and is managed under rain-sheltered plastic greenhouse cultivation conditions. Irrigation, fertilization, pruning, pest and disease management, and vine vigor control were carried out in accordance with local extension recommendations and standard commercial viticultural practices commonly adopted in grape production systems (Du et al. 2015; Thomidis et al. 2016; Reta et al. 2025). Nodal segments were excised from stem nodes containing an axillary bud, and each segment was uniformly trimmed to a length of less than 0.5 cm. To eliminate surface microbial contaminants, the explants were pretreated by immersion in 70% (v/v) ethanol for 30 seconds, followed by surface sterilization in a 1% (w/v) sodium hypochlorite solution for 15 minutes. Subsequently, the explants were rinsed three times with sterile distilled water under aseptic conditions in a laminar airflow cabinet to remove any residual disinfectant. The sterilized explants were cultured according to a previously reported in vitro propagation protocol for ‘Shine Muscat’ (Kim et al. 2023) on the Murashige and Skoog (MS) basal medium, supplemented with 3% (w/v) sucrose and 0.8% (w/v) agar, with the pH adjusted to 5.8 before autoclaving. Stable in vitro cultures were established and maintained under controlled growth conditions (25 ± 1°C, 16/8 h light/dark photoperiod, and a light intensity level of 40 µmol·m-2·s-1 provided by LED lighting). The initial virus infection status of the in vitro plantlets was confirmed by means of reverse transcription–polymerase chain reaction (RT-PCR) testing. The forward and reverse primer sequences used for the detection of GLRaV-3 and GFkV are listed in Table 1. Total RNA was extracted from leaf tissues, and virus-specific primers were employed to detect Grapevine leafroll-associated virus 3 (GLRaV-3) and Grapevine fleck virus (GFkV). Only plantlets that tested positive for both viruses were selected for the subsequent experiments.
Table 1.
Primer sequences and PCR conditions used for the detection of GLRaV-3 and GFkV
| Virus | Primer | Sequence (5’-3’) | Amplicon size (bp) | Cycle |
Annealing (°C) | Reference |
| GLRaV-3 | F | TCT TAA ART AYG TTA AGG ACG G | 340 | 33 | 53 | Kim et al. 2021 |
| R | GGC TCG TTA ATA ACT TTC GG | 340 | 33 | 53 | Kim et al. 2021 | |
| GFkV | F | GTC CTC GGC CCA GTG AAA AAG | 300 | 33 | 53 | Kim et al. 2021 |
| R | CAG GTT GTA GTC GGT GTT GTC | 300 | 33 | 53 | Kim et al. 2021 |
In vitro culture establishment and experimental design
Established V. vinifera L. ‘Shine Muscat’ in vitro plantlets were pre-cultured under identical controlled growth conditions and then divided into four treatment groups: an untreated control group, a thermotherapy group, a ribavirin chemotherapy group, and a group that received the combined thermotherapy–ribavirin treatment. A total of 30 plantlets were used per treatment group. The detailed treatment conditions, including the ribavirin concentration, thermotherapy temperature, treatment duration, and recovery period, are summarized in Table 2. All in vitro cultures were maintained at 25 ± 1°C with a 16/8 h light/dark photoperiod, providing irradiance of 40 µmol·m-2·s-1 using LED lighting. The ribavirin treatment was performed by supplementing the MS medium with 20 or 40 mg·L-1 ribavirin and culturing the plantlets for four weeks. The selected concentration range was based on levels commonly applied in previous in vitro grapevine sanitation studies (10–50 mg·L-1). Earlier reports have demonstrated that ribavirin at 20–40 mg·L-1 effectively suppresses viral RNA synthesis without severely impairing the regeneration capacity (Wang et al. 2018; Zhang et al. 2024). Accordingly, 20 and 40 mg·L-1 were selected in this study for a comparative evaluation of the balance between antiviral efficacy and growth maintenance. Thermotherapy was performed in a controlled growth chamber (Vision Scientific Co., Ltd., Korea; model VS-91G09M-4D), where the temperature was consistently maintained at 37°C throughout the treatment. For the combined treatments, in vitro plantlets on a ribavirin-containing medium were also exposed to 37°C for the same duration. After completing all treatments, the plantlets were transferred to a ribavirin-free MS medium and allowed to recover for two weeks, after which they were used for the subsequent shoot-tip culture experiments.
Table 2.
Summary of sanitation treatments applied to GLRaV-3 and GFkV co-infected ‘Shine Muscat’ plantlets
Thermotherapy and ribavirin treatments
Thermotherapy was applied to whole in vitro plantlets through continuous exposure to a temperature of 37°C for four weeks (28 days), with identical day and night temperatures maintained to minimize physiological stress caused by temperature fluctuations. The selected treatment duration was based on previous in vitro thermotherapy studies reporting that exposure to temperatures around 37°C for approximately three to five weeks (21–35 days) would effectively suppress viral replication while maintaining plant survival and regeneration capacity (Wang et al. 2018). The ribavirin treatment involved culturing plantlets on a MS medium supplemented with ribavirin for four weeks, during which plant growth and physiological status were continuously monitored. For the combined thermotherapy–ribavirin treatments, plantlets cultured on a ribavirin-supplemented MS medium were simultaneously exposed to 37°C for four weeks. After completing all treatments, the plantlets were transferred to fresh MS media without ribavirin and allowed to recover for two weeks. Following this recovery period, shoot apices were excised and subjected to shoot-tip culturing for subsequent regeneration and virus elimination analyses.
Shoot-tip culture and plant regeneration
After each treatment, shoot apices were excised from in-vitro-grown plants under a stereomicroscope. The excised shoot tips measured approximately 0.3 to 0.5 mm and included the apical meristem along with one to two leaf primordia. These shoot tips were immediately transferred to a regeneration medium composed of half-strength MS salts, supplemented with 1.0 mg·L-1 6-benzyladenine (BA) and 0.1 mg·L-1 indole-3-acetic acid (IAA). Cultures were maintained under controlled conditions (25 ± 1°C, 16/8 h light/dark photoperiod, 40 µmol·m-2·s-1). Regeneration and survival of the shoot tips were evaluated after six weeks of culturing. Regenerated plantlets with normal shoot development were then transferred to a multiplication medium for further growth and stabilization prior to the virus detection assays. Virus detection was performed in triplicate for each regenerated plantlet to ensure analytical reliability. The survival rate, regeneration rate, shoot length, and leaf number were recorded to assess the effects of the different treatments on shoot-tip viability and regeneration performance.
Data collection and statistical analysis
All growth traits, regeneration performance outcomes, and virus elimination data were recorded six weeks after the completion of each treatment and are presented as the mean ± standard deviation (SD). Statistical analyses were conducted using Python (version 3.12; Python Software Foundation, USA). Differences among treatments were evaluated by means of a one-way analysis of variance (ANOVA). When significant effects were detected, mean separation was performed using Duncan’s multiple range test at p < 0.05. A principal component analysis (PCA) was conducted to examine multivariate patterns of growth responses following standardization of the variables. The relationship between shoot-tip survival and plant survival was assessed via a Pearson’s correlation analysis and by simple linear regression. The coefficient of determination (R²) was derived from the regression model. The level of statistical significance was set to p < 0.05.
Results
Effects of thermotherapy and ribavirin treatments on the in vitro shoot growth of grapevines
The effects of thermotherapy, ribavirin chemotherapy, and the combined treatments on the growth traits of in vitro ‘Shine Muscat’ plantlets are presented in Table 3 and Fig. 1. Table 3 summarizes the mean values, standard deviations, and statistical differences among the treatments (p < 0.05), while Fig. 1 illustrates treatment-dependent growth response patterns based on PCA. Significant differences were observed among the treatments (p < 0.05), and each growth trait exhibited distinct responses depending on the treatment applied.
Table 3.
Effects of the thermotherapy, ribavirin chemotherapy, and the combined treatments on the in vitro growth traits of ‘Shine Muscat’ grapevine plantlets
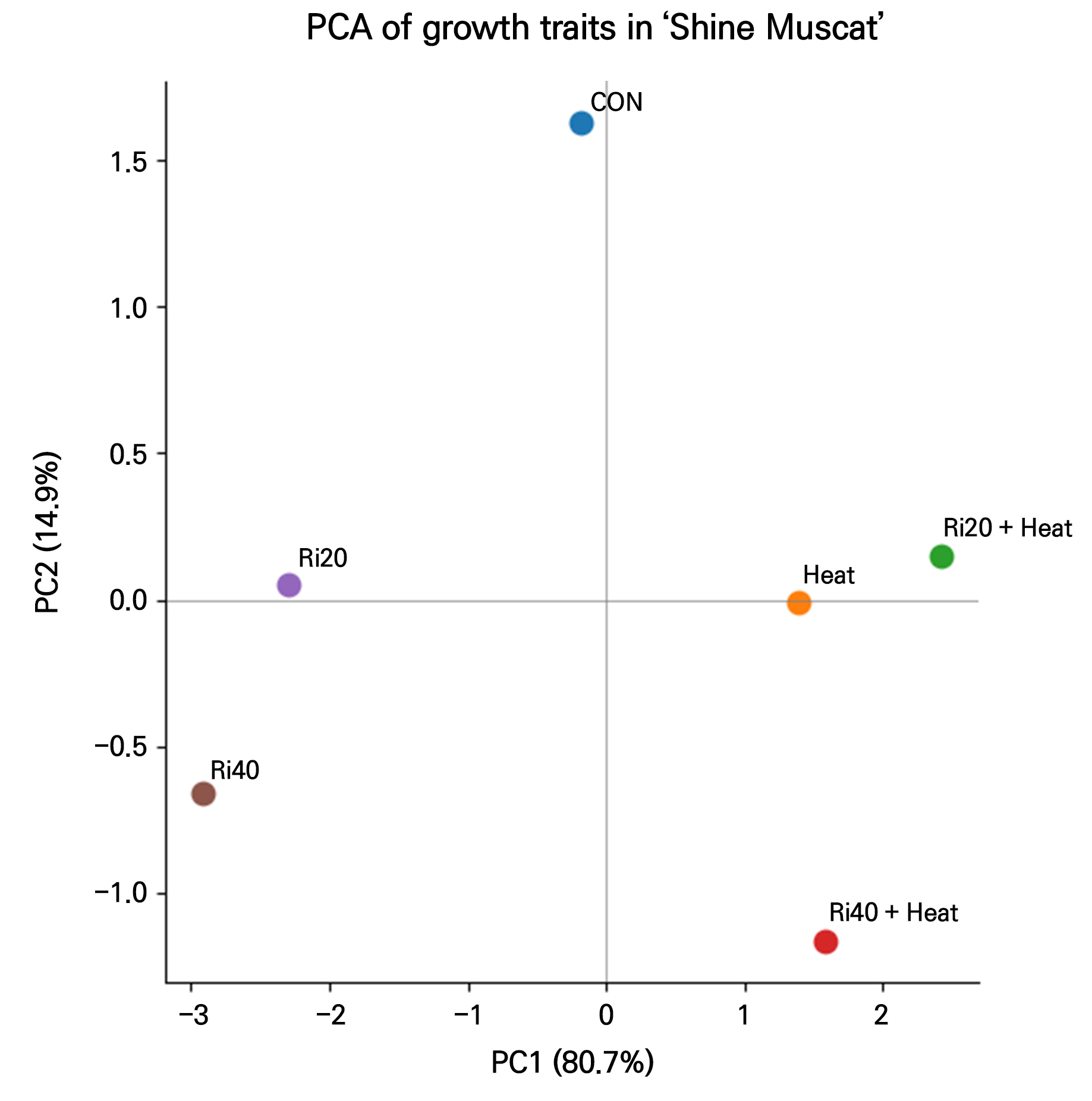
Fig. 1.
Principal component analysis (PCA) of growth traits in in vitro ‘Shine Muscat’ grapevine plantlets under the thermotherapy, ribavirin chemotherapy, and combined treatments. PC1 and PC2 explained 80.7% and 14.9% of the total variance, respectively, showing clear treatment-dependent separation of growth responses.
For plant height, the thermotherapy-only treatment (T) showed the highest value (0.86 cm), which was significantly greater than those of the R20 and R40 treatments. Among the combined treatments, R20+T (0.79 cm) did not differ significantly from T but was significantly higher than the ribavirin-only treatments. In contrast, the R40 treatment exhibited the lowest plant height (0.44 cm), confirming a concentration-dependent inhibitory effect of ribavirin on shoot growth. A similar trend was observed for the number of leaves. The T and R20+T treatments showed relatively high values (2.16 and 2.34 leaves, respectively) compared to R40 (1.81 leaves), with significant differences among the treatments. Comparable patterns were also detected for the numbers of buds and nodes. Notably, the R20+T treatment recorded the highest values for both buds (2.38) and nodes (2.38), which were significantly greater than those observed in the ribavirin-only treatments (R20 and R40). Regarding root length, the untreated control (CON) exhibited the highest value (1.74 cm), whereas R40+T showed the lowest value (0.33 cm), representing a statistically significant reduction. The T and R20+T treatments displayed intermediate values. Overall, the ribavirin-only treatments (R20 and R40) consistently showed lower mean values across all growth traits, indicating a dose-dependent inhibitory effect on in vitro shoot growth. However, the combined treatments (R20+T and R40+T) resulted in significantly higher values for plant height, leaf number, bud number, and node number compared to the ribavirin-only treatments, suggesting that the growth inhibition induced by ribavirin was partially alleviated when combined with thermotherapy.
The PCA revealed clear separation among the treatments in terms of growth responses (Fig. 1). The first principal component (PC1) accounted for 80.7% of the total variance and represented the primary axis of variation among treatments. The ribavirin-only treatments (Ri20 and Ri40) were positioned in the negative direction of PC1, with R40 exhibiting the most negative score, indicating the strongest growth suppression pattern among the treatments. In contrast, the thermotherapy-only treatment (T) and the combined treatments (Ri20+T and Ri40+T) were located in the positive direction of PC1 and were clearly separated from the ribavirin-only treatments. This distribution indicates that the thermotherapy and combined treatments exhibited growth response patterns distinct from those observed under ribavirin alone. The second principal component (PC2), which explained 14.9% of the total variance, further differentiated treatment-specific responses. The untreated control (CON) was clearly positioned in the positive direction of PC2, whereas the Ri40+T treatment was distinctly separated in the negative direction along PC2. This pattern suggests that the high-intensity combined treatment (Ri40+T) produced a unique growth response compared to the other treatments. Overall, the PCA clearly distinguished the ribavirin-only treatments from the thermotherapy and combined treatments, with the Ri40+T treatment exhibiting the most pronounced separation among all treatments.
Changes in shoot-tip regeneration and plant survival under single and combined treatments
The relationship between shoot-tip survival and subsequent plant survival is shown in Fig. 2. The shoot survival rate (%) and plant survival rate (%) for each treatment were plotted in a scatter diagram, and a linear regression analysis was conducted to assess the association between the two variables.
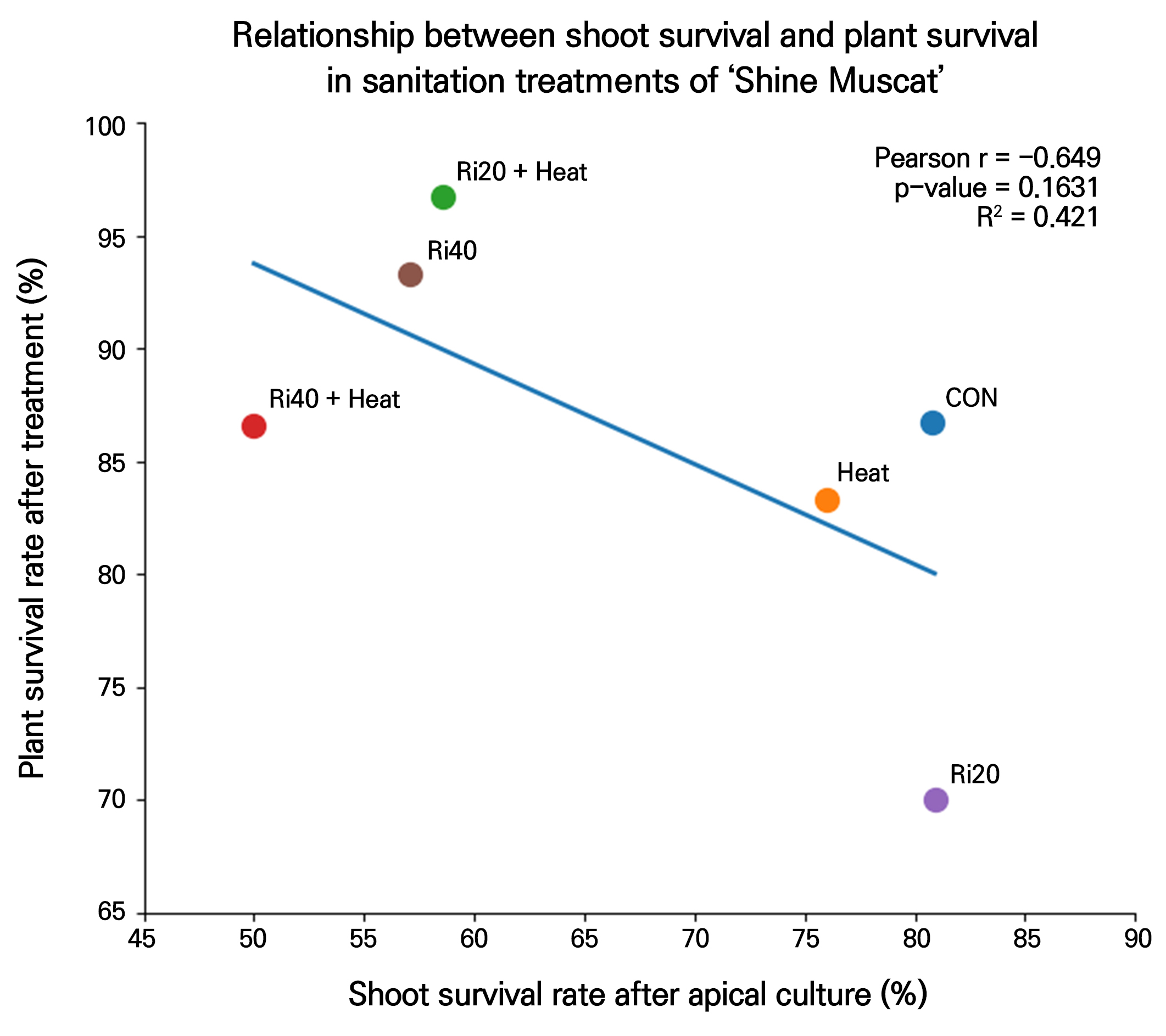
A Pearson correlation analysis indicated a negative trend between shoot-tip survival and plant survival (r = –0.649); however, this correlation was not statistically significant (p = 0.1631). The coefficient of determination (R²) was 0.421, suggesting that shoot-tip survival accounted for approximately 42.1% of the variation in plant survival, although this was insufficient to establish statistical significance.
With respect to the treatment distribution, the untreated control (CON) exhibited both high shoot survival and relatively high plant survival. Among the ribavirin-only treatments, Ri20 showed high shoot survival but relatively low plant survival, whereas Ri40 exhibited intermediate shoot survival and comparatively high plant survival. The thermotherapy-only treatment (T) was positioned at an intermediate level for both parameters. Among the combined treatments, Ri20+T exhibited the highest plant survival despite relatively lower shoot survival, whereas Ri40+T showed reduced values for both parameters.
Overall, although contrasting patterns between shoot-tip survival and plant survival were observed under certain treatment conditions, the relationship between the two variables was not statistically significant. Therefore, these findings are interpreted as treatment-dependent trends rather than evidence of a definitive trade-off.
Comparison of GLRaV-3 and GFkV elimination efficiency rates under different treatment conditions
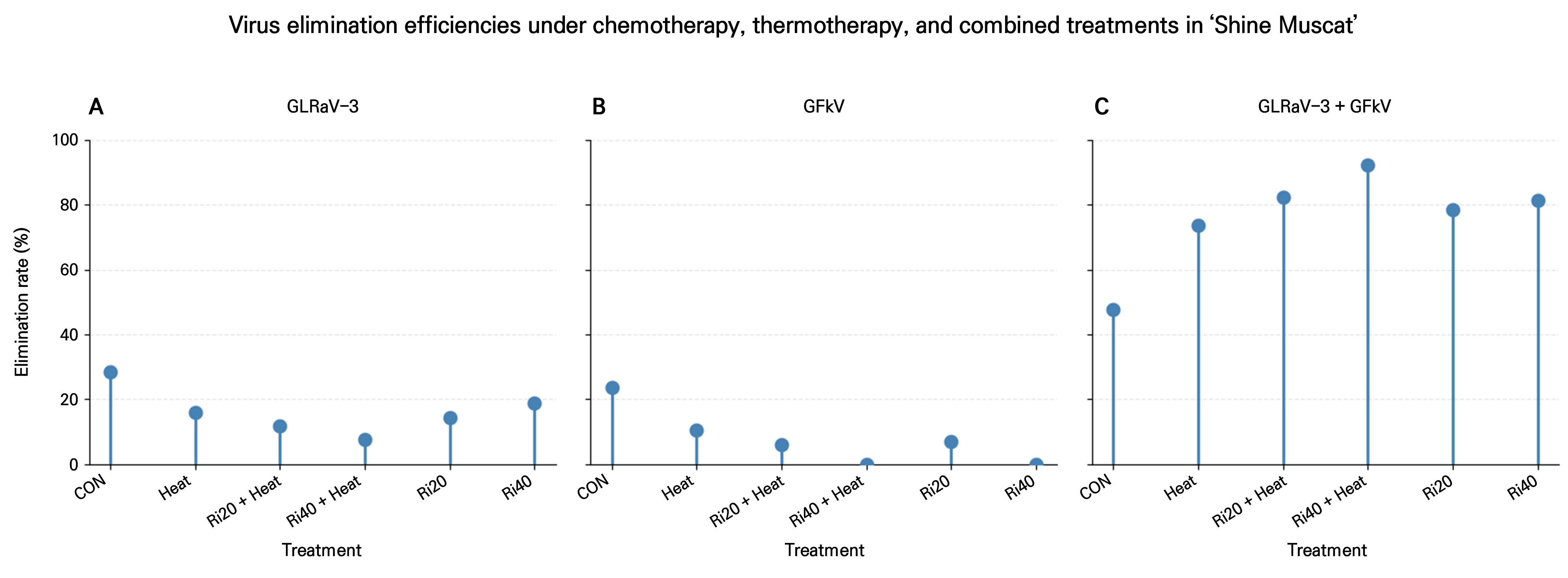
To evaluate the effects of chemotherapy (ribavirin), thermotherapy, and their combined application on virus elimination, we compared the elimination efficiency rates of GLRaV-3, GFkV, and the mixed infection across different treatments (Fig. 3). Three-panel lollipop plots were devised (Fig. 3A–3C), indicating that the elimination efficiency rates of individual viruses were generally low, regardless of the treatment conditions. The elimination rate of GLRaV-3 was 28.6% in the untreated control (CON), while thermotherapy alone (T) and ribavirin treatments (Ri20, Ri40) resulted in limited elimination efficiency rates ranging from 7.7% to 18.8% (Fig. 3A). Elimination of GFkV was consistently lower than that of GLRaV-3, with virus elimination rarely detected in the Ri40 and Ri40+T treatments (Fig. 3B). In contrast, the elimination of the combined infection of GLRaV-3 and GFkV exhibited a markedly different pattern from those of the individual viruses (Fig. 3C). Thermotherapy alone achieved a relatively high mixed-infection elimination rate of 73.7%, while the combined ribavirin–thermotherapy treatments resulted in the highest efficiency rates, reaching 82.4% in Ri20+T and 92.3% in Ri40+T. Notably, the Ri40+T treatment demonstrated the highest elimination efficiency among all treatments for mixed infection.
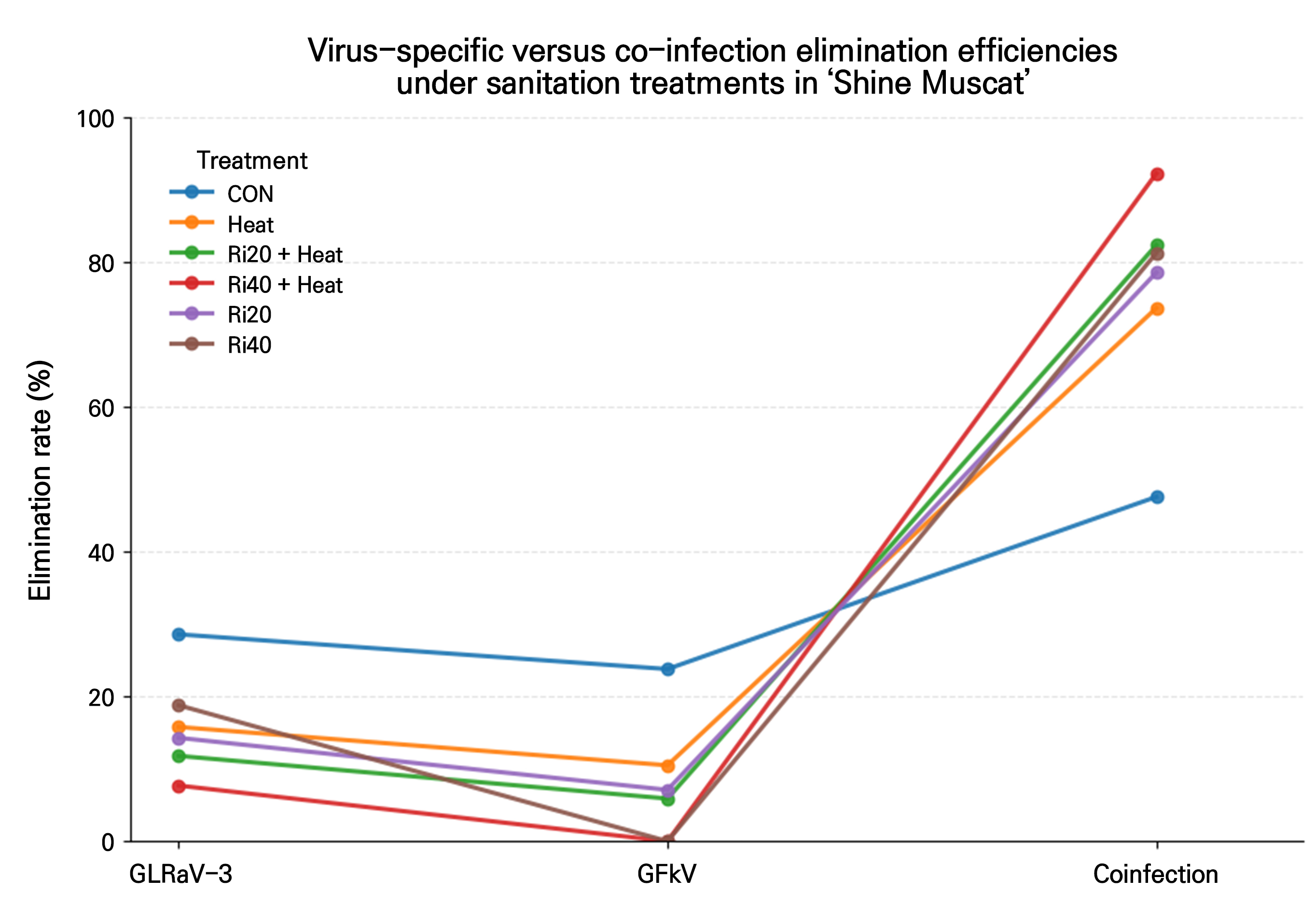
A slopegraph analysis (Fig. 4) clearly illustrated the treatment-specific transitions from individual virus elimination to mixed-infection elimination. Across all treatments, the elimination efficiency rates for GLRaV-3 and GFkV remained low when considered individually, while mixed-infection elimination showed a substantial improvement under the same conditions. Notably, the Ri40+T treatment exhibited the lowest elimination rates for both GLRaV-3 and GFkV individually yet achieved the highest efficiency for mixed-infection elimination. This highlights a structural difference in virus elimination responses between the single and mixed infections, suggesting that treatment effects should be interpreted separately for individual viruses and mixed infections.
Overall, the thermotherapy, ribavirin chemotherapy, and the combined treatments exerted differential effects on growth traits and post-regeneration survival patterns of in vitro ‘Shine Muscat’ plantlets. As shown in Table 3, the ribavirin-only treatments (R20 and R40) exhibited significantly lower values (p < 0.05) for most aboveground growth traits, specifically the plant height, number of leaves, number of buds, and number of nodes, indicating a concentration-dependent decline in growth performance. In contrast, the thermotherapy-only treatment (T) and the combined treatments (R20+T and R40+T) showed relatively high values with regard to aboveground growth traits compared to the ribavirin-only treatments. Root length was longest in the untreated control (CON) and shortest in the R40+T treatment, with significant differences observed among the treatments (p < 0.05). The principal component analysis (Fig. 1) clearly distinguished among the treatments based on the growth responses. PC1, which accounted for 80.7% of the total variance, served as the primary axis separating the ribavirin-only treatments from the thermotherapy and combined treatments, with R40 positioned at the most negative end of the axis. PC2 (14.9% of the total variance) further differentiated the Ri40+T treatment from the other treatments, indicating an additional dimension of variation in the growth response. As shown in Fig. 2, an analysis of the relationship between shoot survival and plant survival revealed a negative correlation trend (r = –0.649); however, this relationship was not statistically significant (p = 0.1631; R² = 0.421). The distribution of treatments suggests that the combination of these two survival parameters varied depending on the treatment type and intensity. Taken together, growth performance, multivariate response patterns, and post-regeneration survival varied according to the treatment type and intensity, underscoring the importance of an integrated evaluation of treatment effects on growth maintenance and regeneration stability.
In contrast, the virus elimination efficiency did not exhibit a simple linear decline with an increase in the treatment stress (Figs. 3 and 4). While the elimination of GLRaV-3 and GFkV individually remained limited across most treatments, the elimination of mixed infections increased sharply under the combined ribavirin–thermotherapy conditions. Notably, the Ri40+T treatment demonstrated the lowest individual virus elimination rate but the highest mixed-infection elimination rate, suggesting that the efficacy of virus elimination under combined treatments may be non-linearly enhanced in cases of mixed infections. These findings emphasize that sanitation efficiency evaluations should extend beyond individual virus elimination and incorporate any existing mixed-infection status. For the practical production of virus-free plants, optimal treatment conditions should be determined by balancing elimination efficiency with regeneration capacity rather than focusing solely on maximizing elimination.
Discussion
In the present study, the comparative responses among treatment groups were clearly differentiated. Ribavirin-only treatments caused the strongest reduction in aboveground growth, indicating that chemotherapy alone imposed the greatest growth penalty. In contrast, thermotherapy alone maintained relatively stable growth performance, suggesting that the applied heat regime was less detrimental to vegetative development. The combined thermotherapy–ribavirin treatments showed intermediate but treatment-dependent responses: R20+T provided a more favorable balance between growth maintenance and sanitation efficiency, whereas R40+T achieved the highest mixed-infection elimination efficiency but was accompanied by greater reductions in growth and post-regeneration survival stability. These comparisons indicate that the effectiveness of sanitation treatments in ‘Shine Muscat’ should be interpreted not only in terms of virus elimination but also in relation to the degree of growth preservation and regeneration stability.
Ribavirin-only treatments resulted in significant reductions in most aboveground growth traits, indicating a concentration-dependent inhibitory effect on plant growth. These findings suggest that the action of ribavirin is not limited to suppression of viral RNA synthesis but may also affect nucleic acid metabolism in host cells (Crotty et al. 2001; Beaucourt and Vignuzzi 2014). Ribavirin is known to act as a guanosine analog and to exert antiviral effects through multiple mechanisms, including depletion of the intracellular GTP pool via the inhibition of inosine monophosphate dehydrogenase, interference with viral RNA polymerase activity, and, in some cases, lethal mutagenesis. However, these effects may also impose metabolic constraints on host tissues, particularly on the cell division and redifferentiation processes required for shoot regeneration under in vitro conditions, thereby contributing to growth inhibition and reduced regenerative performance. In contrast, thermotherapy alone maintained relatively stable growth performance, suggesting that the applied heat regime suppressed viral replication and cell-to-cell movement without critically impairing the viability of meristematic tissues (Vivek and Modgil 2018; Thanuja et al. 2025). Previous studies have shown that thermotherapy can reduce virus distributions within shoot tips, enlarge the virus-free region around the apical dome, and promote viral RNA degradation. Therefore, the relative stability observed under thermotherapy alone in the present study may reflect a more selective antiviral effect with less of a direct metabolic burden on host growth than the ribavirin-only treatment (Graci and Cameron 2006; Wang et al. 2008).
Interestingly, the two combined treatments showed distinct responses depending on the treatment intensity, indicating the presence of a physiological threshold in the host response. The R20+T treatment appeared to provide sufficient antiviral pressure while remaining within a range that still allowed tissue regeneration and growth maintenance (Senula et al. 1999; Ebrahimi et al. 2022; Szabó et al. 2024). In contrast, the R40+T treatment likely imposed a substantially greater burden on host metabolism because the inhibitory effects of the high ribavirin concentration were superimposed on thermal stress, thereby further weakening the regeneration competence of meristematic tissues. This suggests that the effect of combined treatment is not simply additive; rather, it may shift from a complementary interaction at moderate intensity, where improved sanitation efficiency is achieved with acceptable growth preservation, to an excessive stress condition at a higher intensity level, where growth inhibition and the regeneration decline become more pronounced. Such an interpretation is consistent with previous reports describing phytotoxic effects and regeneration impairment associated with intensive in vitro virus elimination treatments (Magyar-Tábori et al. 2021).
The analysis of the relationship between shoot-tip survival and plant survival revealed a negative correlation trend, though this relationship was not statistically significant. This suggests that the two survival indicators do not change in a strictly proportional or oppositional manner across treatment intensity levels (Bettoni et al. 2022; Nongsiang et al. 2025). Rather than representing a definitive structural trade-off, the observed patterns are more appropriately interpreted as treatment-dependent variations in survival responses.
Virus elimination efficiency exhibited patterns that differed from those observed for growth responses. While the elimination rates of individual viruses (GLRaV-3 and GFkV) remained generally low across most treatments, the mixed-infection elimination rate increased markedly under combined thermotherapy–ribavirin conditions. Notably, the Ri40+T treatment showed the lowest individual virus elimination rates yet achieved the highest mixed-infection elimination efficiency. This indicates that elimination under co-infection conditions cannot be explained by a simple additive effect of single-virus removal (Crotty et al. 2001; Karimpour et al. 2025).
The distinct pattern observed for mixed-infection elimination suggests that the co-infected state is not merely defined as the coexistence of two independent viruses but is rather a biologically interconnected condition involving host physiological responses and virus–virus interactions. GLRaV-3 infection has been reported to reduce photosynthesis, stomatal conductance, electron transport, and shoot growth, thereby weakening overall host physiology. Under a mixed infection, such effects may be further amplified through oxidative imbalance and more severe physiological disruption (Hančević et al. 2022; Segredo-Otero and Sanjuán 2022). Therefore, the high mixed-infection elimination efficiency achieved under the combined treatment may not reflect the simple independent removal of each virus but rather the destabilization of the physiological and molecular interaction network that sustains the co-infected state (Endeshaw et al. 2014; Andika et al. 2023). This interpretation further supports the conclusion of the present study that sanitation efficiency cannot be adequately explained by single-virus elimination rates alone, particularly in systems where mixed infections are prevalent.
These findings suggest that combined thermotherapy and ribavirin treatments may destabilize the biological interaction network of co-infecting viruses rather than selectively eliminating individual viruses. Co-infection systems may exhibit distinct biological dynamics compared with single-virus infections, and combined treatments may exert non-linear effects on these interactions (Segredo-Otero and Sanjuán 2022; Andika et al. 2023). Consequently, sanitation assessments based solely on single-virus elimination rates may underestimate the actual sanitation performance achieved under co-infection conditions.
Taken together, this study provides three key implications. First, increasing the treatment intensity differentially affects growth maintenance and regeneration stability. Second, the relationship between shoot-tip survival and plant survival cannot be generalized as a fixed trade-off structure. Third, mixed-infection elimination efficiency may follow response patterns that are independent of single-virus elimination rates.
Therefore, grapevine sanitation strategies should move beyond the exclusive maximization of single-virus elimination and adopt an integrated evaluation framework that simultaneously considers growth preservation, regeneration stability, and mixed-infection elimination efficiency. In cultivars where mixed infections are prevalent, incorporating the co-infection elimination efficiency rate as a core assessment metric may provide a more practical basis for the production of stable virus-free planting materials.
Conclusion
This study shows that the combined use of ribavirin chemotherapy and thermotherapy can effectively improve the elimination of co-infections in V. vinifera ‘Shine Muscat’ while maintaining in vitro growth and regeneration stability. Among the treatments tested, Ri20+T achieved the best balance between shoot-tip regeneration stability and virus elimination efficiency, making it an optimal choice for producing virus-free plants. These findings underscore the significance of sanitation strategies that combine efficient co-infection elimination with regeneration potential, offering a rational framework for nursery-level sanitation that extends beyond traditional evaluations focused solely on single-virus infections.
