

서 언
재료 및 방법
꽃가루받이용 4배체 선발
2배체 씨 없는 수박 종피특징 및 품질 비교
4배체 수박 꽃가루 특성조사
결과 및 고찰
꽃가루받이용 4배체 선발
2배체 씨 없는 수박 종피 특징 및 품질 비교
4배체 수박 꽃가루 특성
서 언
수박은 영양학 및 경제적으로 중요한 원예작물 중 하나이다(Kombo and Sari, 2019). 국내 수박 생산액은 7,946억원으로 채소류 중에서는 5번째, 과채류에서는 딸기 13,057억원 다음으로 비중이 높은 작물이다(MAFRA, 2017b). 수박 재배면적과 생산량은 1995년까지 증가하다가 점차 감소하는 추세에 있다. 2016년 기준 재배면적은 13,440ha로 1995년 대비 70.3% 감소했고, 생산량은 570,160톤으로 49.1% 감소하였다(MAFRA, 2017a). 재배면적 대비 생산량의 감소가 상대적으로 적었던 이유는 품종개량, 재배기술 향상과 시설재배 비율 증가 때문으로 여겨진다. 이처럼 수박 전체 생산과 소비는 줄어들고 있지만 3배체 씨 없는 수박은 2000년대 초반부터 본격적으로 재배가 시작된 이후 지속적으로 재배면적이 늘어나는 추세이다. 일반 2배체 수박은 고온기에 접어들면 시들음증과 육질악변과(피수박) 발생이 증가하며 야간에 호흡량 증가로 당도가 떨어진다(Kano, 2004). 반면 3배체 씨 없는 수박은 초세가 강건하고 생육적온이 일반수박보다 높아 고온기에도 상대적으로 고품질 수박을 생산할 수 있다. 이와 같은 이유로 고온기에 3배체 씨 없는 수박을 재배하는 농가가 증가하고 있으며 더불어 당도가 높고 먹기가 편하다는 장점 때문에 씨 없는 수박에 대한 소비자들의 선호도도 높아지고 있다(Marr and Gast, 1991; Phat et al., 2017). 하지만 3배체 씨 없는 수박은 발아율이 낮아 종자 가격이 비싸고 기상여건이 좋지 않을때 착과율이 떨어지며 공동과와 같은 비상품과 발생이 많고 중심부와 주변부의 당도 차이가 크다는 단점 등이 대두되고 있다(Phat et al., 2015; Diezma-Iglesias et al., 2004).
3배체 품종을 이용해서 씨 없는 수박을 생산하는 방법 외에도 생장조절제, 연 X-ray처리 꽃가루, 4배체 꽃가루를 이용하면 씨 없는 수박을 생산할 수 있다(Hayata and Niimi, 1995; Sugiyama and Morishita, 2000; DB HiTek Inc., 2001; Pandolfini, 2009). 이중 실용화된 기술은 연 X-ray 처리 꽃가루를 이용하는 방식으로 꽃가루에 연 X-ray를 조사해서 수분 시 종자가 발달되지 않도록 한다. 연 X-ray를 조사한 꽃가루를 암술머리에 인공수분 시켜주면 꽃가루관 신장부터 수정까지는 정상적으로 이루어지지만 수정된 배가 분화되지 못하고 퇴화되어 종자가 형성되지 않는다(Sugiyama and Morishita, 2002). 하지만 연 X-ray처리 꽃가루를 이용하는 방법은 꽃가루의 활력을 떨어뜨리지 않고 수집 및 저장할 수 있는 기술이 뒷받침되어야 하며 연 X-ray를 처리하기 위해서는 장비와 전문인력이 필요해 비용이 많이 든다.
X-ray 처리 꽃가루 대신 4배체 수박의 꽃가루를 이용해도 2배체 씨 없는 수박을 생산할 수 있다. 4배체 수박의 암꽃에 2배체 수박의 꽃가루를 인공수분 시켜주면 3배체 종자가 발달되지만 반대로 2배체 수박 암꽃에 4배체 수박 꽃가루를 인공수분 시켜주면 배는 발달하지 않고 미숙종자의 종피만 남으며 자방은 정상적으로 발달하여 씨 없는 수박이 된다. 4배체 수박 꽃가루를 이용해서 씨 없는 수박을 생산하는 방식에 대한 연구는 거의 이루어지지 않아 아직 실용화되지 못했다. 본 연구는 연 X-ray 처리 꽃가루를 이용해서 2배체 씨 없는 수박을 생산하는 방식을 4배체 수박 꽃가루를 이용하여 생산하는 방식으로 대체할 수 있는지 그 가능성을 검토하고 해당 기술을 발전시킬 수 있는 방안을 찾기 위해 수행되었다.
재료 및 방법
꽃가루받이용 4배체 선발
전라북도농업기술원 수박시험장에서 육성 및 수집한 4배체 6계통 WC-8C-2C, GW11090031, GW11090032, 14SS3-1, 14SS3-2, 14SS3-3에 대해 계통별 개화 수꽃 수, 자가수분 후 착과율과 종자 수량을 조사하였다. 각 계통별 10주씩 정식 후 본엽이 5매 전개됐을 때 줄기의 선단 생장점을 잘라 분지를 유도하였으며 그 중 1차분지 3줄기만 남기고 제거하였다. 남겨진 3줄기를 제외한 추가 1차분지와 2차분지는 지속적으로 제거하였다. 개화 수꽃 수 조사를 위해 가장 먼저 개화한 2번째 암꽃 개화일로부터 2주간 개화한 총 수꽃의 수를 집계하여 1주당 개화한 수꽃의 수를 계산하였다. 각 4배체 수박 계통들의 착과율과 종자수량은 3번째 암꽃에 인공수분(자가수분)하여 착과시킨 수박을 이용하여 산출하였다.
인공수분은 암꽃 1개당 수꽃 1개를 사용하였으며 수꽃의 꽃잎을 제거한 후 암꽃의 주두에 꽃가루가 충분히 묻도록 가볍게 문질러서 진행하였다. 4배체 계통들과 함께 2배체 ‘조생삼복꿀’ 수박을 정식하였고, 각 4배체 계통의 수꽃으로 35주의 ‘조생삼복꿀’ 수박에 인공수분하여 착과된 2배체 씨 없는 수박의 착과율과 과실 품질을 조사하였다. 당도는 과실의 과경 접합부위(꼭지)에서 화흔부(배꼽)까지 세로축을 따라 2등분 후 중앙부에서 과즙을 내어 디지털당도계(PAL-1, ATAGO Inc., Japan)를 사용하여 측정하였다. 4배체 계통별 종자량과 계통별 꽃가루를 2배체 암꽃에 인공수분하여 착과된 수박의 착과율과의 관계 분석을 위해 엑셀(Excel 2016, Microsoft Inc., USA)의 데이터 분석 기능을 이용하여 통계분석을 하였다.
2배체 씨 없는 수박 종피특징 및 품질 비교
2배체 씨 없는 수박의 착과율과 과품질은 4배체 수분수로 선발한 GW11090032 계통의 수꽃을 ‘조생삼복꿀’ 수박의 암꽃에 인공수분하여 착과시킨 수박을 대상으로 조사하였다. 인공수분은 2배체 암꽃 1개당 4배체 수꽃 1개를 사용하여 진행하였다. 더불어 연 X-ray 처리된 SWT(Seedless Watermelon Technology, OREC Inc., Japan) 꽃가루를 ‘조생삼복꿀’ 수박에 인공수분하여 생산한 2배체 씨 없는 수박과 ‘조생삼복꿀’ 수박을 자가수분해서 생산한 일반 수박의 과실 품질과 종피 특징을 조사하여 비교하였다.
4배체 수박 꽃가루 특성조사
4배체 GW11090032계통과 2배체 ‘조생삼복꿀’의 꽃가루관 발아율과 형태적 특징을 조사하였다. 꽃가루관의 발아 특성은 배지를 이용하는 방법(in vitro)과 인공수분 시킨 암술을 적출하여 염색 후 관찰하는 방법(in vivo)으로 조사하였다. 꽃가루관 발아시험용 배지는 Brewbaker and Kwack(1963)의 방법에 따라 증류수 1L에 sucrose 100g, boric acid 0.1g, calcium nitrate 0.3g, magnesium sulfate 0.2g, potassium nitrate 0.1g을 넣고 pH를 5.7로 조정한 후 agar 10g을 넣고 멸균하여 사용하였다. 개화 당일 오전 10시경에 수집한 꽃가루를 배지에 치상하여 30°C로 설정한 배양기에서 1시간 동안 발아시켜 꽃가루관의 발아율을 조사하였다. 꽃가루관 발아율은 실체현미경을 25배율로 조정한 배지의 4개 지점을 촬영하여 전체 꽃가루와 발아된 꽃가루를 집계해 계산하였다. 암술 내부의 꽃가루관 관찰은 개화 당일 인공수분한 암꽃을 24시간 경과 후에 채취하여 FAA (formalin acetic acid alcohol) 용액에서 12시간 고정시켰다. 고정된 시료를 60°C의 1N NaOH에서 중탕하여 연화 시킨 후 aniline blue 0.1% 용액에 24시간 침지시켜 염색했다. 염색된 시료는 형광현미경을 이용해 관찰하였다.
꽃가루 형태 관찰은 개화 당일 오전 10시경 수집한 꽃가루의 원상태와 배지에서 올려 수분을 흡수했을 때의 꽃가루, 그리고 초산분해 후의 꽃가루를 관찰하였다. 초산분해는 먼저 꽃에서 수술을 수집한 후 아세트산과 황산을 9:1의 비율로 섞은 혼합용액에 넣고 water bath에서 80°C로 15분간 중탕하였다. 중탕한 시료는 원심분리 후 상징액은 버리고 다시 아세트산을 넣고 tube mixer로 혼합하였다. 이를 0.1mm 표준체로 거른 후 아세트산을 넣고 원심분리 시킨 후 상징액을 제거하였다. 다시 증류수를 넣고 잘 섞어 원심분리 후 상징액을 제거하였다. 여기에 5% KOH를 넣고 water bath에서 5분간 가열한 후 원심분리시켜 상징액을 제거하였다. 25% ethyl alcohol을 넣고 원심분리한 후 상징액을 따라냈다. 남아있는 꽃가루는 건조시킨 후 광학현미경(Eclipse 50i, Nikon, Tokyo, Japan)과 전자현미경(S-2460N, Hitachi, Tokyo, Japan)으로 관찰하였다.
결과 및 고찰
꽃가루받이용 4배체 선발
4배체 6계통 중 2주간 개화한 수꽃이 가장 많았던 계통은 14SS3-3으로 수박 1개체당 평균 19.8개였다. 반면 수꽃이 가장 적었던 계통은 WC-8C-2C로 14SS3-3의 절반 수준으로 4배체 계통에 따라 개화하는 수꽃의 수에 차이가 있음을 확 인 할 수 있었다. 자가수분에 의해 착과된 수박 개수도 계통에 따라 차이가 컸다. GW11090031과 14SS3-1은 10개체 중 1개체에서만 착과가 이루어졌고 GW11090032과 14SS3-3은 각각 6개와 7개가 착과되었다. 수박 과실 1개에 들어있는 종자는 GW11090032가 87개로 다른 계통들과 비교해 월등히 많았고 14SS3-1, 14SS3-2, 14SS3-3은 종자량이 20-30개 정도였다. 이는 4배체 계통 간에 종자량 차이가 많았다고 한 Jaskani et al.(2005)의 보고와 같았다(Table 1).
Table 1. Number of bloomed male flower, fruits and seeds of each tetraploid watermelon line
각 4배체 계통의 수꽃으로 2배체 수박 암꽃에 인공수분한 결과 착과유도가 가장 잘 된 계통은 GW11090032으로 80%의 착과유도율을 보였다. 나머지 계통을 이용했을 때는 50 - 60% 정도의 착과율을 보였다. 꽃가루 제공 계통에 따른 수확기의 2배체 씨 없는 수박 과중과 당도는 차이를 보이지 않았다(Table 2).
Table 2. Fruit set rate and weight of diploid seedless watermelon plants pollinated from each tetraploid watermelon line
yEach value represents mean ± SD.
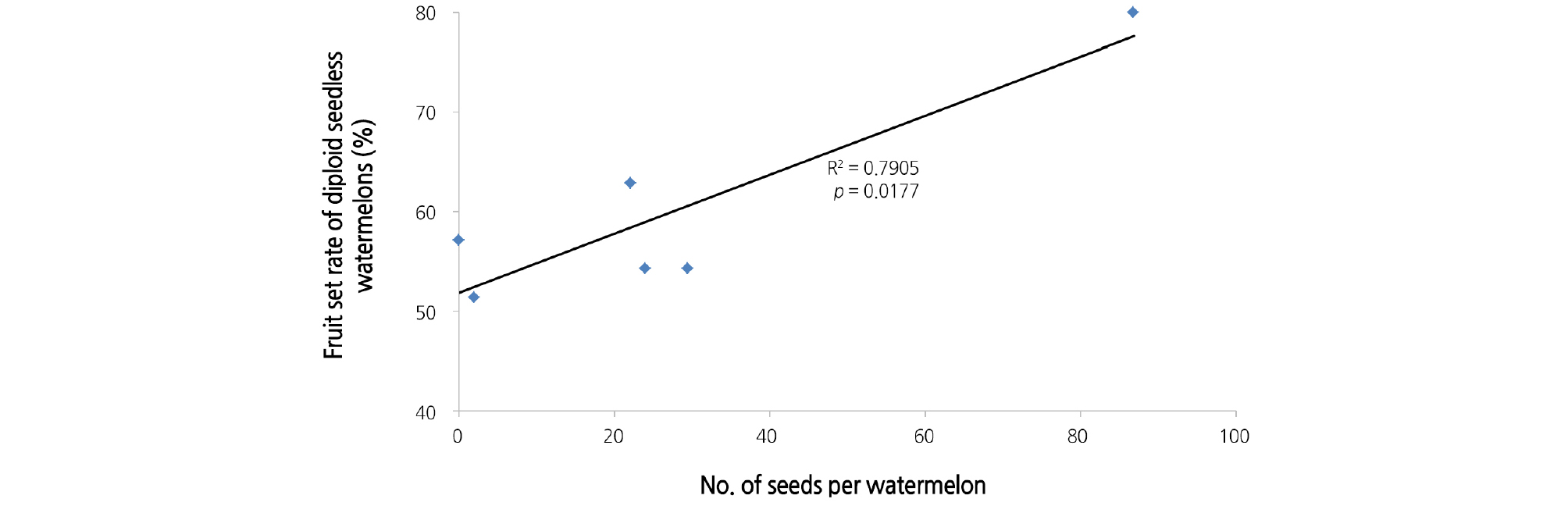
4배체 수박 계통별 자가수분을 통해 발달 및 채종된 종자량과 4배체 계통별 수꽃을 이용해 착과시킨 2배체 씨 없는 수박 착과율과의 관계를 분석한 결과 상관관계가 높게 나타났다(R2 = 0.790, p < 0.05)(Fig. 1). 4배체 각 계통마다 꽃가루의 발아 및 수정능력이 달라 계통에 따라 자가수분시의 채종량과 2배체 수박 암꽃에 인공수분했을 때의 착과율이 달랐던 것으로 판단된다. 올리브(Methamem et al., 2015)와 목화(Kakani et al., 2005)에서도 품종에 따라 꽃가루의 수정능력이 다르다는 보고가 있다. 따라서 2배체 씨 없는 수박 생산을 위한 꽃가루받이용 4배체 선발에 자가수분시의 채종량이 유용한 지표로 활용할 수 있을 것으로 생각된다. 본 시험에서는 채종량이 많았고 2배체 씨 없는 수박의 착과 유도가 잘 됐던 GW11090032 계통을 꽃가루받이용으로 선발하여 다음 실험에 활용하였다.
2배체 씨 없는 수박 종피 특징 및 품질 비교
선발된 4배체 계통 GW11090032의 꽃가루와 SWT 꽃가루를 이용해서 생산한 2배체 씨 없는 수박과 ‘조생삼복꿀’ 수박을 자가수분시켜서 생산한 일반 수박의 착과율과 과실 품질을 비교하였다. 착과율은 SWT 꽃가루 인공수분 처리구가 92.3%, ‘조생삼복꿀’ 자가수분 처리구가 91.9%로 유사하였고, 4배체 꽃가루를 인공수분했을 때의 착과율 75.0%보다 높았다. 반면, 과중, 당도, 과피 두께 등의 과실 품질에서는 차이가 없었다(Table 3).
Table 3. Fruit set rate and characteristics of watermelons pollinated with SWT pollen, pollen of tetraploid plants and pollen of diploid plants
yMean separation within columns by Duncan's multiple range test (p ≤ 0.05).
xJSG : Josaengsambokggul cultivar (self-pollinating).
미숙종자의 종피색은 경우 SWT 꽃가루 인공수분 수박에서 백색에 가까웠지만 4배체 수박 꽃가루 인공수분 수박에서는 연한 갈색 이었다. SWT 꽃가루 인공수분 수박의 일부에서 검정색 종피가 발견되었지만 배는 포함하고 있지 않았다. 미숙종자 100개의 무게는 4배체 수박 꽃가루 인공수분 수박에서 좀더 무거웠으나 길이의 차이는 없었다(Table 4). 결과적으로 4배체 수박 꽃가루로 인공수분해서 생산한 수박은 일반 2배체 수박과 비교해 완숙 종자가 없고, 착과율이 낮다는 점을 제외하고 품질면에서는 차이가 없었다.
Table 4. Seed or seed coat characteristics of watermelons pollinated with SWT pollen, pollen of tetraploid plants and pollen of diploid plants
| Pollenz | Length (mm) | Weight of 100 seed coats (g) | Photo of seed coats |
| SWT pollen (soft X-ray irradiated) | 7.59 ± 1.57y | 3.53 |  |
| GW11090032 (tetraploid) | 7.91 ± 1.21 | 3.80 |  |
| JSGx (diploid) | 10.29 ± 0.63 | 10.16 |  |
yEach value represents mean ± SD.
xJSG : Josaengsambokggul cultivar (self-pollinating).
4배체 수박 꽃가루 특성
4배체 수박의 꽃가루로 인공수분 했을 때 착과율이 낮았던 원인을 찾기 위해 꽃가루관 발아율을 조사한 결과 in vitro 발아실험에서의 화분관 발아율은 2배체 수박의 꽃가루는 29.2%, 4배체 수박의 꽃가루는 14.5%로 2배체 수박의 꽃가루가 4배체 수박의 꽃가루 보다 약 2배정도 높았다(Fig. 2A, B). 이는 멜론에서 배수체 식물체 꽃가루의 발아율이 2배체보다 낮았다는 보고(Sus´ın and Álvarez, 1997)와 유사한 결과이다. In vivo 발아실험에서 암술대를 통과하는 꽃가루관은 2배체 수박의 꽃가루가 4배체 수박의 꽃가루 보다 월등히 많았다(Fig. 2C, D).
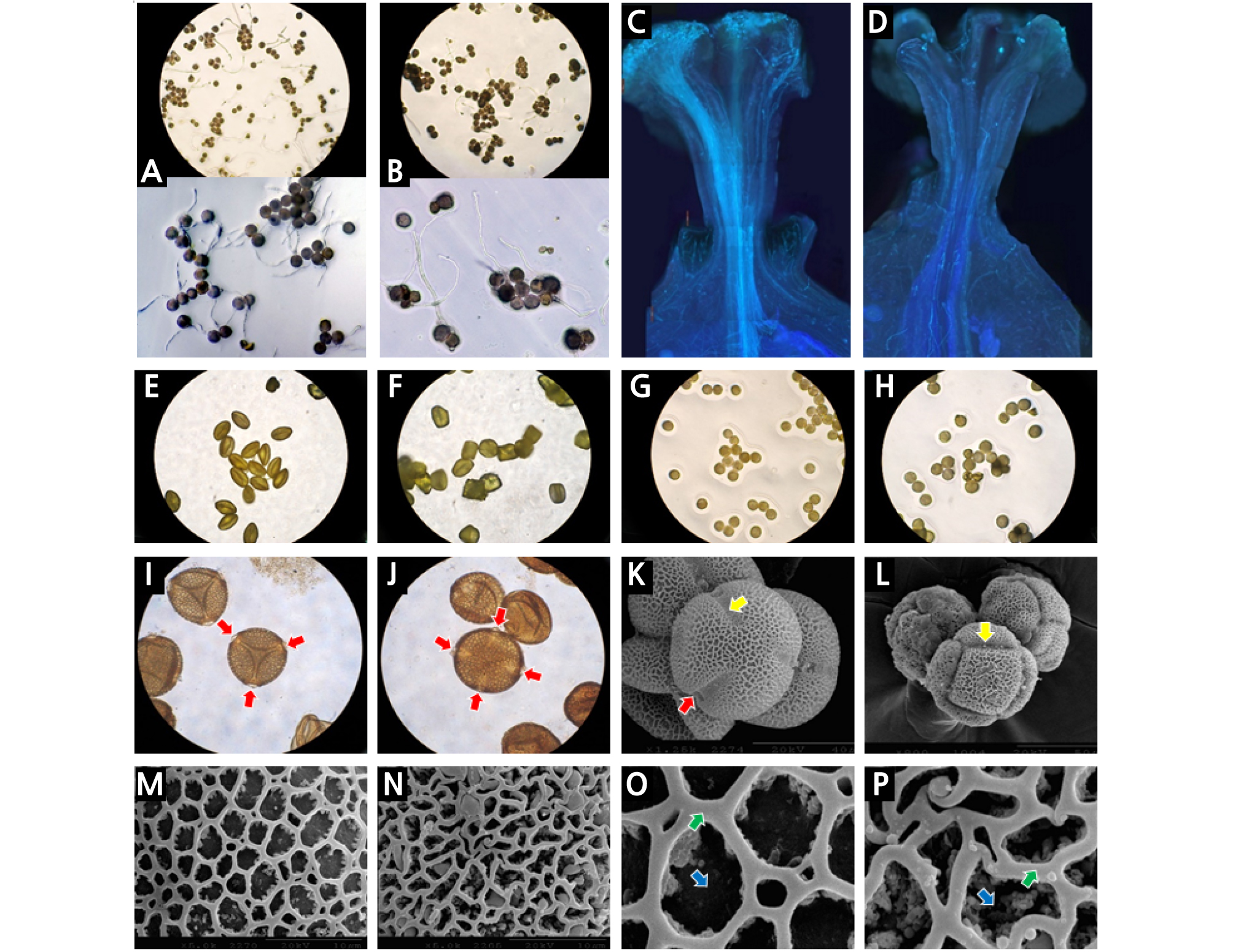
Fig. 2.
Comparison of diploid and tetraploid watermelon pollens: extension of pollen tube in vitro (A) and in vivo (C) of diploid, in vitro (B) and in vivo (D) of tetraploid watermelon pollens (pollens and pollen tubes are light blue color in the picture); diploid watermelon pollens before (E) and after (G) water absorption; tetraploid watermelon pollens before (F) and after (H) water absorption; diploid (I) and tetraploid (J) watermelon pollens after acetolysis (pore: red arrow); scanning electron microscope image (colpus: yellow arrow, pore: red arrow) of diploid (K) and tetraploid (L) watermelon pollens; pollen wall structure (muri: blue arrow, lumina: green arrow) of diploid (M and O) and tetraploid (N and P) watermelon pollens.
4배체 수박의 꽃가루와 2배체 수박의 꽃가루의 형태를 비교하기 위해 꽃가루를 광학현미경으로 관찰한 결과 2배체 수박의 꽃가루는 비교적 균일한 방추형의 낱알로 구성되어 있었으나 4배체 수박의 꽃가루는 각각의 낱알들의 형태가 일정하지 않았다(Fig. 2E, F). 꽃가루가 수분을 흡수했을 때의 2배체 수박 꽃가루는 낱알들이 구형의 일정한 형태로 변했으나 4배체 수박 꽃가루는 크기가 균일하지 않았고 구형으로 팽창하지 못해서 불완전한 형태의 낱알들이 발견되었다(Fig. 2G, H). 2배체 수박 꽃가루는 화분관구는 삼공구형으로 3개의 발아구와 발아공이 같이 존재했다(Fig. 2I, K). 표면은 망상형태를 띄며 부정형의 망강이 표면에 분포하고 망벽은 원주상의 기둥으로 되어있다(Fig. 2M, O). 4배체 수박의 꽃가루는 발아구와 발아공의 유무가 각 꽃가루마다 달랐으며 그중 일부에서는 발아공이 4개인 꽃가루가 관찰되었다(Fig. 2J, L). 표면의 망상 무늬는 균일하지 않았고 망강의 폭은 좁았으며 찌그러진 형태를 보였다(Fig. 2N, P).
꽃가루 특성을 조사한 결과, 착과율이 낮은 원인은 4배체 수박 꽃가루의 꽃가루관 발아율이 낮았기 때문이며, 이는 꽃가루의 불완전한 형태와 발아공의 부재 등 꽃가루 형성과정에서 발달이 정상적이지 못하였기 때문으로 판단된다. 배수성 천남성의 꽃가루모세포가 감수분열이 비정상적으로 일어나 임성이 낮고 결실률이 낮았다는 Liu et al.(2012)의 보고와 유사한 결과이다. 위와 같은 사실을 기초로 볼 때 꽃가루의 발달이 정상적으로 이루어지고 그에 따라 꽃가루의 활력이 높아 착과 유도가 잘 되는 4배체 계통을 육성한다면 연 X-ray 처리 꽃가루를 대체해서 2배체 씨 없는 수박 생산에 활용할 수 있을 것으로 보인다.
