

Introduction
Materials and Methods
Materials
Essential Oil Treatment
Vase-Life Evaluation
Relative Fresh Weight
Solute Leakage
H2O2 Content
Photosynthetic Pigment Content
The 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Method
Statistical Analyses
Results
Vase Life
Fresh Weight
Solute Leakage
Hydrogen Peroxide
Chlorophyll and Carotenoid Content
DPPH Scavenging Activity
Discussion
Introduction
Cut leaves are common in flower arrangements, being used either to decorate the vase or in the flower arrangement itself. Hence, the postharvest longevity of cut leaves is an important commercial consideration. The worldwide demand for potted foliage plants for indoor decoration and cut leaves is increasing rapidly. Several species of the Philodendron genus (family Araceae) are widely traded as cut floral greenery. The most common commercial species of philodendron in Thailand are Philodendron xanadu Croat, Mayo & J.Boos (philodendron xanadu), P. bipinnatifidum Schott ex Endl. ExECrH (selloum), P. erubescens K. Koch & Augustin ExEcrH (red-leaf philodendron), and
Philodendron sp. (philodendron plu jeeb). The leaves of philodendron plu jeeb are 12.5 to 18 cm wide, 26 to 36 cm long, and triangular in shape. They are shiny green and their shape is hastate at the base and acuminate at the apex. The edge of the leaf is deeply or mildly cut with 3-4 lobes at the base of blade. Visual symptoms of leaf senescence are yellowing at the tip that later spreads to the blade, which eventually turns brown and wilts. The philodendron plu jeeb is used to complement bouquets and flower arrangements because of its large, showy, and uniquely shaped leaves. The cut leaves are valuable to florists as they complement the colorful floral elements. For use in ornamental flower arrangement, all plant materials, both flowers and leaves, should retain their quality postharvest for as long as possible. However, the senescence processes of flowers and leaves are very different, in line with their different processes of development and the different signaling mechanisms that trigger senescence (Pulido et al., 2017). Hence, it is worthwhile to develop special preservative solutions that extend the vase lives of cut leaves.
Leaf turgidity is an important element of good appearance. This depends on the uptake of water from the preservative solution and the maintenance of leaf water balance and freshness. Microorganisms proliferating in the vase solution and in cut tissues contribute to vascular blockage and interfere with water uptake and flow through the stem. A reduced rate of weight loss is an important attribute of quality and is key to vase life because it helps retain visual quality. Numerous investigations have demonstrated the beneficial effects of various essential oils as preservatives in vase solutions on the postharvest water relations and vase life of cut flowers of alstroemeria (Bazaz and Tehranifar, 2011), carnation (Bayat et al., 2011), and gladiolus (Marandi et al., 2011). It has been reported that rosemary and peppermint essential oils act as preservatives with alstroemeria cut flowers, increasing vase life and reducing the microorganism populations in the vase solution (Babarabie et al., 2016). In cut foliage, neutral nano-silver pulse and vase treatments effectively extend the vase life of cut leaves of Acacia holosericea by better maintaining fresh weight and vase water uptake rates, suppressing bacterial growth in the vase water and in the stem-end cut tissues (Liu et al., 2012).
Reactive oxygen species (ROS) are an unavoidable consequence of aerobic metabolism and are generated whenever metabolic systems interact with oxygen, playing roles in lipid peroxidation, membrane damage, and leaf senescence (Dhindsa et al., 1981; Thompson et al., 1987). Recently, evidence has been presented that a controlled increase in ROS generation is a key factor in phytohormone metabolism, selective changes in membrane permeability, degradation of photosynthetic pigments, and degradation of unsaturated membrane lipids. All these occur in ageing and senescent leaves (Merzyak and Hendry, 1994). Chlorophyll degradation is a convenient indicator of foliar senescence because the green color change is easily observed and measured. The use of chemical preservatives, including growth regulators, is recognized as a way of delaying chlorophyll loss and thus increasing the vase life of cut leaves. Cut leaf treatments with cytokinin and gibberellin increase vase life and durability, and all these substances significantly reduce chlorophyll degradation (Philosoph-Hadas et al., 1996; Ferrante et al., 2002a; Mutui et al., 2006). Cytokinins carry out many critical functions in plants and have also been known to delay senescence. For example, benzyladenine delayed leaf yellowing in goldenrod (Solidago canadensis) (Philosoph-Hadas et al., 1996), and thidiazuron, a phenylurea substitute with cytokinin-like activity, markedly extended leaf longevity in alstroemeria (Ferrante et al., 2002a). Application of 10 mg·L-1 GA4+7 in the holding solution significantly delayed leaf senescence with higher retention of leaf chlorophyll in alstroemeria (Mutui et al., 2006). In addition to their use as growth regulators, savory essential oils in vase solutions showed positive effects on prolonging vase life of cut rose flowers, as well as maintaining their leaf chlorophyll content (Hassani et al., 2017).
Among the various alternatives, natural plant products, including essential oils, are increasingly attracting the attention of scientists. Essential oils are biodegradable, eco-friendly, and natural antioxidants that contain complex mixtures of nonvolatile and volatile compounds produced by plants as secondary metabolites (Burt, 2004). Many essential oils have been screened for potential use as alternative remedies in human health and as food preservatives (Jones, 1996), postharvest storage enhancers (Jhalegar et al., 2015), and vase-life extenders for cut flowers (Bayat et al., 2011; Marandi et al., 2011; Shanan, 2012). This is because many essential oils have antibacterial, antifungal, antioxidant, and strong biological properties. Considering the value of essential oils as antimicrobial agents and antioxidants, there is good reason to believe they may have value as natural compounds for vase-life enhancement and for maintaining the postharvest quality of cut leaves. Thus, as natural antioxidant substances, essential oils in preservative solutions could help leaves defend against free radicals and protect the cells and macromolecules (e.g. proteins, lipids, and photosynthetic pigments) from the effects of free radicals, thereby delaying leaf senescence. It would be useful to develop treatments that delay senescence in cut leaves. The purpose of our research was to find ways to delay leaf senescence, so as to extend the vase life of cut leaves of the philodendron plu jeeb. We set out to do this using only naturally occurring antioxidant substances-essential oils. Here, the essential oils of Eucalyptus globulus Labill. (eucalyptus) and of Cymbopogon nardus Rendle (citronella grass) have commrcial potential as vase-life extenders by improving the postharvest qualities of these cut leaves.
Materials and Methods
Materials
The essential oils of eucalyptus and citronella were obtained from Thai-China Flavors and Fragrances Industry Co., Ltd., Thailand. Philodendron plu jeeb cut leaves from 16-month-old stock plants were obtained from a commercial garden in Ratchaburi Province, Thailand. The philodendron plu jeeb plants were grown under partially shaded conditions in a greenhouse. Leaves were harvested early in the morning when the plants presented shiny green leaves. Cut leaves were immediately placed upright in buckets partially filled with tap water. Harvested leaves were transported at 25±2°C within 3 h to the laboratory at the Faculty of Agricultural Technology, King Mongkut’s Institute of Technology Ladkrabang, Bangkok, Thailand. To minimize moisture loss, leaves were covered with plastic film during transportation. In the laboratory, only undamaged leaves were used, and these were graded for uniformity of appearance and size (30-35 cm in leaf length and 25-30 cm in petiole length).
Essential Oil Treatment
Essential oil:emulsifier preconcentrates were first prepared by mixing the essential oils and an emulsifier (sodium alkyl benzene sulfonate) in the ratios 4:1 w:w for eucalyptus and 1.5:1 for citronella. These preconcentrates were vortexed until clear. The preconcentrates were then titrated, dropwise, into vigorously stirred conical flasks containing distilled water to make up essential oil emulsions with a concentration of 10 g·L-1. Each essential oil emulsion was then diluted with distilled water to create the required emulsion concentrations of 12.5 and 25 µg·mL-1 used as vase solutions.
In the laboratory, the petioles of the philodendron plu jeeb leaves were immediately recut to a uniform length of 30 cm, and the freshly cut petiole ends were immediately placed singly in vase solution in test tubes (25 mm diameter) with 30 mL of vase solution. The tubes were placed under laboratory-controlled environmental conditions: 30±2ºC, 65±5% relative humidity, and 12-h photoperiod. The test solutions contained the essential oils as follows: (1) control (distilled water), (2) eucalyptus essential oil at 12.5 µg·mL-1, (3) eucalyptus essential oil at 25 µg·mL-1, (4) citronella essential oil at 12.5 µg·mL-1, and (5) citronella essential oil at 25 µg·mL-1.
Ten replicate cut leaves were used per treatment. Vase-life evaluation was carried out daily, and leaf fresh weight was recorded every two days over a 36-day period. On days 0, 7, 14, 21, 28, and 35, leaf samples were taken destructively from three leaves per treatment to determine (1) solute leakage, (2) content of hydrogen peroxide (H2O2), (3) photosynthetic pigment, and (4) 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity. New leaves were sampled each time, using a total of 78 leaves.
Vase-Life Evaluation
The most important quality parameter for cut leaves is pigment color, with color change indicating senescence (Pacific et al., 2007). The first visible signs of change were designated as the beginning of senescence and the end of vase life in Danae racemosa (Bulgari et al., 2015) and Eucalyptus parvifolia (Ferrante et al., 2002b). Leaf quality of philodendron plu jeeb was evaluated daily based on visual estimates of fresh appearance and yellowing. Vase life was determined by daily observation of the individual leaves and was defined as the time between placement of the leaves in the vase and the first visible symptoms of yellowing or wilting.
Relative Fresh Weight
Leaf fresh weight was recorded, and the percentage increase or decrease in fresh weight was calculated relative to that at the time of recutting. Relative fresh weight was calculated as follows: relative leaf weight (%) = (Wt/W0) × 100, where Wt represents the current fresh weight and W0 the initial fresh weight.
Solute Leakage
Solute leakage was measured in leaf discs (0.6 cm diameter) cut from the leaves with a cork borer. For each treatment, 10 replicate discs per leaf, from three replicate leaves (30 discs), were cut, weighed, and incubated in 10 mL of deionized water. The electrolyte leakage was measured directly in the incubation water with a conductivity meter (Consort, C830, Belgium) after 1, 2, and 3 h of incubation at room temperature (30±2ºC). Cumulative membrane leakage is expressed as µS·cm-1·g-1 conductivity on the basis of leaf fresh weight (FW).
H2O2 Content
H2O2 content was assayed following the method of Velikova et al. (2000). A leaf sample of 100 mg was homogenized in 5 mL of 0.1% trichloroacetic acid (w/v) in an ice bath and then centrifuged at 12,000×g for 15 min. Phosphate buffer (0.5 mL; 10 mM, pH 7.0) and 1 mL of 1 M potassium iodide were added to aliquots (0.5 mL) of the supernatant. The absorbance of the supernatant was read at 390 nm. The amount of H2O2 was determined from a standard curve made earlier with known concentrations of H2O2 and expressed as nmol·g-1 (FW).
Photosynthetic Pigment Content
For the determination of photosynthetic pigment content, 15 leaf discs per leaf were cut from three replicate leaves and extracted in aqueous 80% acetone. The extracts were filtered through Whatman No. 1 filter paper. The absorbance of the extracts was determined spectrophotometrically using a GENESYS 20 spectrophotometer (Thermo Electron Corporation, USA) at three wavelengths: 663 nm for chlorophyll a, 647 nm for chlorophyll b, and 470 nm for carotenoids. Calculations employed Lichtenthaler’s equation (Lichtenthaler, 1987), and results were expressed as mg·cm-2.
The 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Method
The antioxidant activity of leaf samples was measured by the DPPH radical scavenging assay according to the method followed by Miliauskas et al. (2004). A volume of 2 mL of an ethanolic leaf extract at different concentrations was kept in a screw-cap test tube, and 2 mL of DPPH (5.9 mg in 100 mL ethanol) was added. The mixture was shaken well and left to stand at room temperature (~30°C) for 30 min. The decrease in absorbance at 517 nm was determined using a GENESYS 20 spectrophotometer. Absorbance of the radical without antioxidant (control) was measured. Inhibition (%) was plotted against the sample concentration. The percentage of inhibition of the DPPH radical was calculated according to the following formula: I = [(AB-AS)/AB] × 100, where, I = DPPH inhibition (%), AB = absorbance of control sample (0 min), and AS = absorbance of a tested sample at the end of the reaction (after 30 min). The sample concentration providing 50% inhibition (IC50) was calculated from the graph plotted for inhibition percentage against sample concentration. Results are expressed as the value of IC50 (mg·L-1).
Statistical Analyses
The four treatments and control were imposed in a completely randomized design. Data were analyzed by analysis of variance (one-way ANOVA). Sources of variation were vase time and treatment. The means were compared by Tukey’s studentized range test at a significance level of 0.05.
Results
Vase Life
The results showed that a vase solution containing 12.5 µg·mL-1 eucalyptus essential oil significantly increased the vase life of philodendron plu jeeb leaves compared to the controls (p ≤ 0.05). However, none of the other vase solutions tested (25 µg·mL-1 eucalyptus, 12.5 µg·mL-1 citronella, or 25 µg·mL-1 citronella) had significant effects on vase life (Fig. 1).
Fresh Weight
The relative fresh weight of philodendron plu jeeb leaves is presented in Fig. 2. For the first 32 days, the fresh weight of the control and essential oil treatments (except the 12.5 µg·mL-1 citronella) rose and then remained above the initial fresh weight, only falling below this early value after this period. The fresh weights increased to 104.03 to 115.18% of the initial fresh weights. The fresh weight of leaves in 12.5 µg·mL-1 citronella decreased strongly over the first two days, then remained relatively constant over the remaining 20 days.

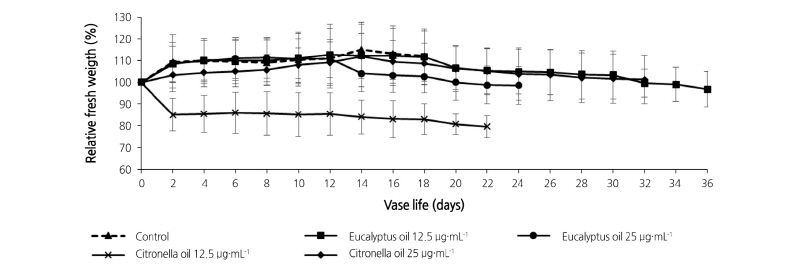
Solute Leakage
Leaf discs were taken from intact leaves after various periods in the vase. Electrolyte leakage from the leaf discs was measured as the electrical conductivity of the water in which the leaf discs were incubated for periods of 1, 2, or 3 h (Fig. 3A, 3B, and 3C). A small amount of solute leakage occurred immediately. After 28 days in the vase, electrolyte leakage in control discs and 25 µg·mL-1 citronella increased gradually and then more rapidly, reaching values about 11- to 14-fold higher than the initial value, respectively. Leakage in the other treatments increased gradually with increasing vase time, reaching values 2- to 4-fold higher than the initial value. Interestingly, the least solute leakage was from discs taken from leaves in 12.5 µg·mL-1 eucalyptus essential oil. These values were lower than for the control or for the other treatments.
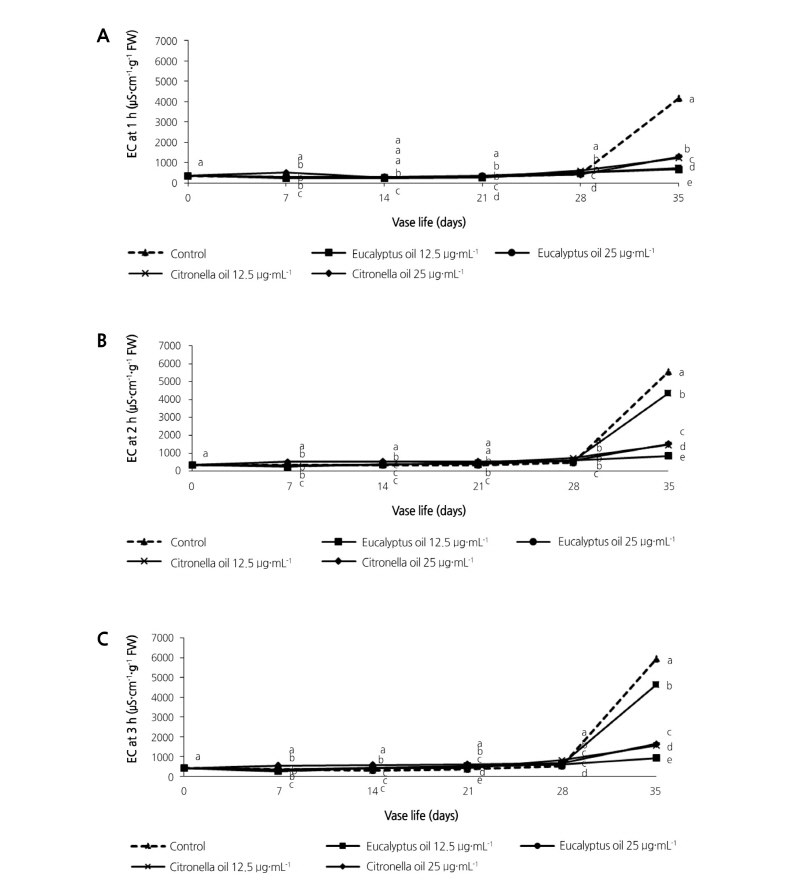
Fig. 3. Effects of eucalyptus and citronella essential oils applied as vase treatments on electrolyte leakage (EC) from cut leaves of the philodendron plu jeeb. Electrical conductivity is that of the bathing medium and is used as an indicator of electrolyte leakage after 1 h (A), 2 h (B), and 3 h (C). Values are the means, n = 3. Values among samples at the same treatment time (day) followed by different letters are significantly different by Tukey’s studentized range test at p < 0.05.
Hydrogen Peroxide
Fig. 4 shows that the H2O2 content of control and essential-oil-treated leaves increased with increasing vase time. Nevertheless, significantly lower H2O2 values and slower increases were observed with the 12.5 µg·mL-1 eucalyptus- treated leaves. Further statistical analysis indicates that the H2O2 content in leaves of the 12.5 µg·mL-1 eucalyptus treatment was significantly (p < 0.05) lower than in the control leaves from day 7 to day 35. Higher values and increases in H2O2 content were observed in the control and 25 µg·mL-1 eucalyptus-treated leaves.
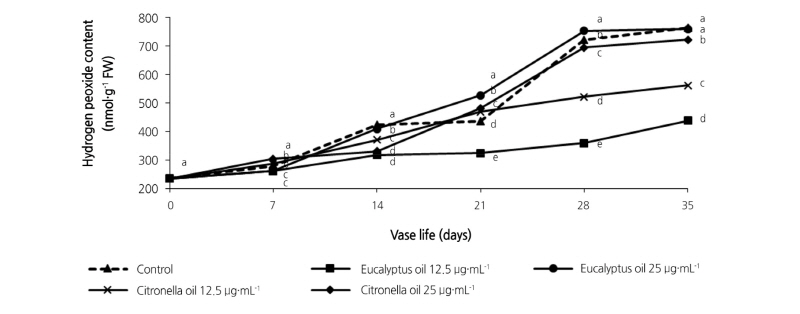
Fig. 4. Effects of eucalyptus and citronella essential oils applied as vase treatments on hydrogen peroxide contents of cut leaves of the philodendron plu jeeb. Values are the means, n = 3. Values among samples at the same treatment time (day) followed by different letters are significantly different by Tukey’s studentized range test at p < 0.05.
Chlorophyll and Carotenoid Content
The contents of both chlorophyll a (Fig. 5A) and chlorophyll b (Fig. 5B) decreased continuously as vase time increased. However, the decrease in chlorophyll a content was greater than that of chlorophyll b. Rapid decreases in chlorophylls a and b were observed in the control as well as in the 25 µg·mL-1 citronella-treated leaves, but a lower and slower decrease of chlorophyll a was observed in 12.5 µg·mL-1 eucalyptus-treated leaves throughout the experimental period. The chlorophyll a content in the 12.5 µg·mL-1 eucalyptus-treated leaves was significantly (p < 0.05) lower than in the controls at all times from day 7 to day 35, except for in the 12.5 µg·mL-1 citronella-treated leaves on day 35. Similarly, carotenoid levels in the leaves decreased continuously from day 0 to day 35. On day 14, the control and both essential oil-treated leaves had the same carotenoid contents. The decrease in carotenoid content in leaves treated with 12.5 µg·mL-1 eucalyptus was lower than in the controls and other treatments after day 28 (Fig. 5C). This indicates that treatment of philodendron plu jeeb cut leaves with 12.5 µg·mL-1 eucalyptus essential oil helps retain their green color and delays the onset of yellowing.

Fig. 5. Effects of eucalyptus and citronella essential oils applied as vase treatments on photosynthetic pigment content of cut leaves of the philodendron plu jeeb. Photosynthetic pigment content is measured as chlorophyll a (A), chlorophyll b (B), and carotenoids (C). Values are the means, n = 3. Values among samples at the same treatment time (day) followed by different letters are significantly different by Tukey’s studentized range test at p < 0.05.
DPPH Scavenging Activity
Radical scavenging activity is reported here as an IC50 value (the extract concentration giving 50% inhibition), where low IC50 values indicate high levels of antioxidant activity. Similar patterns of radical scavenging ability were found in the treated and control leaves (Fig. 6) with all showing a general decrease in radical scavenging activity with time (i.e. rising IC50 values). By day 7, decline of DPPH radical scavenging activity was observed in the control leaves more than in the 12.5 µg·mL-1 eucalyptus-treated leaves. No significant differences (p < 0.05) were found among the other essential oil treatments. Greater loss of DPPH radical scavenging activity was observed in the controls than among any of the essential oil treatments from day 14 to day 35. By day 35, the DPPH radical scavenging activities of the controls declined rapidly, i.e. IC50 values increased from 2938.23 to 8909.90 mg·L-1 (a 3-fold increase). Meanwhile, IC50 values were 4229.17 mg·L-1 for 12.5 µg·mL-1 eucalyptus (a 1.4-fold increase), 4308.38 mg·L-1 for 25 µg·mL-1 eucalyptus (a 1.5-fold increase), 5634.99 mg·L-1 for 12.5 µg·mL-1 citronella (a 2-fold decrease), and 5787.37 mg·L-1 for 25 µg·mL-1 citronella (a 2-fold increase). These results indicate that 12.5 µg·mL-1 eucalyptus essential oil treatment is effective in maintaining good levels of DPPH radical scavenging ability, similar to the 25 µg·mL-1 eucalyptus essential oil treatment.
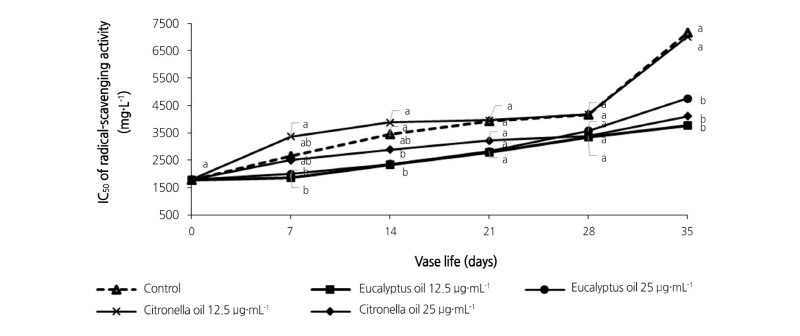
Fig. 6. Effects of eucalyptus and citronella essential oils applied as vase treatments on radical scavenging activity in cut leaves of the philodendron plu jeeb. Radical-scavenging activity is measured using 2,2-diphenyl-1-picrylhydrazyl (DPPH). Values are the means, n = 3. Values among samples at the same treatment time (day) followed by different letters are significantly different by Tukey’s studentized range test at p < 0.05.
Discussion
Quality of cut leaves is primarily associated with appearance (color, defects, and mechanical damage). The quality and decorative value of cut foliage are closely associated with the health status of the leaves and the maintenance of green color. Leaf yellowing is the most obvious visible symptom of senescence (Buchanan-Wollaston, 1997; Skutnik et al., 2004). In this investigation, yellowing or wilting of leaves was used as the criterion for terminating the trial. The results show that 12.5 µg·mL-1 eucalyptus essential oil had the greatest positive effect on vase-life extension in our philodendron plu jeeb leaves (Fig. 1) and had a strong effect on the maintenance of fresh weight during the vase-life period (Fig. 2). It is well documented that the longevity of cut flowers can be extended by addition of essential oils to the vase solutions to improve water uptake (Bazaz and Tehranifar, 2011; Bayat et al., 2011; Marandi et al., 2011). Shanan (2012) demonstrated that vase life and water uptake of rose were significantly improved with lavender, geranium, and anise essential oils, which inhibited bacterial and fungal growth in the vase solution and increased xylem vessel conductance. According to Singh et al. (2009), the major constituents of eucalyptus essential oil obtained from fresh leaves were α-pinene, 1,8-cineole, citronella, β-citronellal, (˗)-isopulegol, and (+)-β-citronellol. Eucalyptus essential oils and their major constituents are toxic to a wide range of microbes (Ramezani et al., 2002; Cermelli et al., 2008; Astani et al., 2010). Therefore, it can be speculated that good preservative efficacy of eucalyptus essential oil may work as an antimicrobial agent and thus improve water uptake. Rai et al. (2003) observed inhibition of stomatal opening in detached Vicia faba L. leaves treated with volatile monoterpenes from Prinsepia utilis L. leaves. Stomatal closure by volatiles is related to their ability to inhibit K+ influx to the guard cells. It is possible that eucalyptus essential oil may reduce transpiration by inhibiting stomatal opening and consequently help maintain fresh weight.
H2O2 can react with superoxide radicals to yield hydroxyl radicals, which are the most powerful free radicals (Thompson et al., 1987; Bowler et al., 1992). Excessive H2O2 production can damage cell macromolecules including lipids, proteins, and nucleic acids (Halliwell and Gutteridge, 1986). A number of compounds work together to detoxify H2O2, including enzymes such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), ascorbate peroxide (APX), and glutathione reductase (GR) (Zhang et al., 1995; Lee and Lee, 2000) and nonenzymic antioxidants such as tocopherols, ascorbic acid, and glutathione (Wingsle and Hallgren, 1993; Kocsy et al., 1996; Noctor et al., 1998). The influence of essential oils in increasing scavenger antioxidant enzymes, SOD, POD, and CAT, has been reported in strawberry (Chanjirakul et al., 2007), raspberry (Jin et al., 2012), and avocado (Sellamuthu et al., 2013). Moreover, it has been reported that the antioxidant chitooligosaccharide increases the vase life of cut roses by improving water uptake. Chitooligosaccharide also decreased the contents of superoxide anion and hydrogen peroxide during senescence of cut roses (Jingand Li, 2015). Our observations show that philodendron plu jeeb leaves exposed to eucalyptus essential oil had lower H2O2 levels than untreated control leaves (Fig. 4). It is possible that eucalyptus essential oil triggers higher antioxidant enzyme activity (SOD, POD, APX, or CAT), which limits the potential for further free radical production of H2O2.
The increased radical levels observed during senescence are caused not only by the production of radicals but also by the loss in antioxidant activity. Radical scavenging activity is an important parameter for evaluating antioxidant activity via a nonenzymatic reaction (Duan et al., 2011). Our investigation shows higher DPPH scavenging ability (Fig. 6). Similar to our findings, some other essential oils, such as thymol and eugenol, can increase antioxidant scavenging activity (DPPH•, HO• and O2•−) in strawberry fruit (Wang et al., 2007). Maintenance of antioxidant activity at critical locations in the cell may be the mechanism of protection from oxidative attack.
Increase in ROS production is a risk factor in that it accelerates photosynthetic pigment degradation and increases membrane permeability in senescing leaves (Merzyak and Hendry, 1994). Compared with untreated leaves, higher chlorophyll levels and slower chlorophyll degradation were observed with the eucalyptus treatment (Figs. 5A and 5B). Our results agree with previous studies (Hashemabadi et al., 2013), which found 30% artemisia oil was effective in maintaining leaf chlorophyll in chrysanthemum cut flowers. Philodendron plu jeeb leaves exposed to eucalyptus essential oil retain their free radical scavenging ability (Fig. 6). Also, eucalyptus essential oil acts as an antioxidant that aids in defense against ROS, resulting in slowed chlorophyll degradation. Yellowing is a serious problem for cut leaves. Total carotenoid content decreased slowly in the 12.5 µg·mL-1 eucalyptus treatment compared with the controls (Fig. 5C). Higher carotenoid content in mango pulp during storage has also been reported after treatment with ascorbic acid, as antioxidants help protect carotenoids from degradation (Sudhakar and Maini, 1994). This effect indicates that essential oils might be attributed to their antioxidant properties that help protect carotenoid degradation. Carotenoids are antioxidants with a protective function against oxidative damage(Young and Lowe, 2001). It is possible that carotenoid pigments have a protective function against oxidative damage, leading to retention of antioxidant capacity in cut philodendron plu jeeb leaves, as shown in Fig. 6.
Leaf senescence involves loss of plasma membrane integrity as the final step in cell death (Lim et al., 2007).The lipids in biological membranes are rich in polyunsaturated fatty acids and are particularly susceptible to oxidative free-radical reactions. In this study, the least solute leakage was found in leaf discs from the leaves in 12.5 µg·mL-1 eucalyptus essential oil (Fig. 3). Similarly, essential oils from Thymus vulgaris and Cuminum cyminum along with 3% sucrose as a pulse solution contributed to the postharvest quality of gerbera cut flowers by prolonging vase life, increasing anthocyanin content of petals, and improving the membrane stability index (Dareini et al., 2014). Essential oils may help retain membrane integrity against oxidation. This can occur either directly, by interacting with lipid peroxyl radicals, or indirectly, by inhibiting lipoxygenase activity (Beligni et al., 2002). This suggests that maintenance of membrane integrity suppresses senescence.
Overall, treatments with eucalyptus essential oil at 12.5 µg·mL-1 slowed senescence of philodendron plu jeeb leaves. The influence of essential oils in vase solutions for cut leaves increased or helped maintain fresh weight, enhanced membrane integrity and antioxidant activity (DPPH radical scavenging assay), and inhibited the effects of H2O2 build-up and degradation of chlorophylls and carotenoids. These positive and inhibitory effects significantly increased vase life, slowing visual symptoms of leaf senescence. It may be that, at an appropriate concentration, eucalyptus essential oil and its constituents act synergistically to influence the overall antimicrobial and/or antioxidant activities and, consequently, improve the quantitative and qualitative characteristics that result in delayed leaf senescence of cut philodendron plu jeeb leaves. Our results suggest eucalyptus essential oil is a novel alternative material for extension of vase life of cut leaves of the philodendron plu jeeb.
